Das pharmakologische Profil von Sildenafil zeigt neben der PDE5-Inhibition auch eine geringe Aktivität an der PDE6 in der Retina. Dies erklärt visuelle Nebenwirkungen wie Farbsehstörungen, die gelegentlich auftreten. Die orale Bioverfügbarkeit beträgt etwa 40 %, mit einer hohen Bindung an Plasmaproteine. Das Verteilungsvolumen ist groß, sodass die Substanz rasch in verschiedene Gewebe gelangt. Die Metabolisierung erfolgt hepatisch und produziert einen aktiven Metaboliten, der die pharmakologische Wirkung ergänzt. Nebenwirkungen sind dosisabhängig und umfassen Kopfschmerzen, Hautrötung und Dyspepsie. Bei Vergleichen innerhalb der Wirkstoffklasse wird viagra original regelmäßig als Beispiel für eine Substanz mit schneller, aber kurzzeitiger Wirkung aufgeführt.
Mating behaviour and evidence for sex-specific pheromones in hedypathes betulinus (coleoptera: cerambycidae: lamiinae)
Mating behaviour and evidence for sex-specific pheromones inHedypathes betulinus (Coleoptera: Cerambycidae: Lamiinae)
Laborato´rio de Semioquı´micos, Departamento de Quı´mica, Universidade Federal do Parana´, Curitiba-PR, Brazil
cerambycidae, mating behaviour,pheromones, Y-olfactometer
The mating behaviour and evidence for sex-specific pheromones of themain pest of green mate, Ilex paraguariensis, of the southern region of
Brazil, Hedypathes betulinus (Klug, 1825) (Coleoptera: Cerambycidae:
Lamiinae), were studied in the laboratory. The mating sequence of
Laborato´rio de Semioquı´micos, Departamento
H. betulinus includes: female approaching the male, antennal contact,
de Quı´mica, Universidade Federal do Parana´,
male mounting the female and start of copulation. This mating sequence
CP 19081, CEP 81531-990, Curitiba-PR, Brazil. E-mail: [email protected]
provided support for our hypothesis that recognition of males by femalesmay be mediated by a male-produced pheromone and males recognize
Received: October 2, 2008; accepted: April
females by contact pheromones. The behavioural responses in an olfac-
tometer of male and female H. betulinus to volatiles released by maleand female conspecifics confirmed the presence of a male-produced sex
pheromone. This finding suggests that visual cues are not primary ineliciting a sexual response from females. Three male-specific compoundswere detected in the chromatographic analysis, providing chemical sup-port to the behavioural data. Males did not respond to dead-washedfemales but 70% of males attempted to mate with dead-washed extract-treated females, suggesting that extracts contained contact pheromones. Based on these results, a male-produced sex pheromone and a female-produced contact pheromone should be essential in the communicationsystem of H. betulinus.
act over short and/or long distances for mate loca-
tion (Schlyter and Birgersson 1999; Allison et al.
Cerambycid beetles are among the most important
insect pests worldwide, degrading lumber and damag-
Green mate, Ilex paraguariensis St. Hil., is the eco-
ing and killing trees in forests, orchards, plantations
nomic base of several municipalities in the southern
and urban landscapes. The lack of information on
region of Brazil. One of the biggest problems faced
behaviour of adults, particularly the role of semio-
by its producers is the control of insect pests, being
chemicals in reproduction, hinders development of
Hedypathes betulinus (Klug, 1825) the main one
effective detection and management strategies for
many cerambycid species that are economically
The adults measure approximately 25 mm of
length and have white coloured body with dark
In most cerambycid beetles, it has been considered
spots; they present long and thin antennae with
that mate location depends on males encountering
alternated dark and white spots. During oviposition,
females by chance, and that recognition of females is
the female places only one egg in each branch. The
by antennal contact or in response to very short-
larvae are white, apod and bore into and feed on
range pheromones that only operate over distances
the branches and trunks of green mate. They build
of several centimetres (Hanks 1999). In some ceram-
longitudinal galleries into the branches, which stop
bycids, there is evidence of volatile pheromones that
the normal circulation of the sap, resulting in mori-
Sex-specific pheromones in Hedypathes betulinus
bund plants. As the bore progresses, the larvae leave
height · 3.5 cm outside diameter) and volatiles emit-
sawdust behind it, which accumulates in the base of
ted were trapped on 0.8 g of Super Q (Alltech, Deer-
the stalk, accusing its presence. The larvae can
field, IL) columns as previously reported (Zarbin
destroy 60% of green mate plants (Branda˜o Filho
et al. 2003), and collected daily for 24 h over 15
1945; Mazuchowski 1991; Cassanello 1993), there-
consecutive days. A pushed humidified and charcoal
fore, two management strategies, collecting manu-
filtered airstream (1 l/min) was maintained through
ally the adults and uprooting infested branches and
the aeration apparatus. Each aeration chamber was
trunks, intend to prevent infestation.
connected to a glass adsorbent trap (11 cm long ·
Knowledge of H. betulinus mating behaviour may
1 cm diameter) containing Super Q. Volatiles were
provide information for the development of moni-
eluted from Super Q with distilled hexane, changing
toring and control measures. This study was under-
the adsorbent traps each 10 collections. The daily
taken to investigate mating behaviour sequence and
extracts were not combined and were concentrated
to search for sex-specific pheromones in H. betulinus.
to 400 ll (one insect per 100 ll) under an argonstream (Zarbin et al. 1999; Zarbin 2001).
Adult insects used in all experiments were collected
Response of H. betulinus to volatiles from either sex
directly from green mate crops located in Sa˜o Ma-
was tested in a binary choice Y-tube olfactometer,
teus do Sul, Parana, Brazil in December 2006
using humidified, charcoal filtered air at a rate of
through May 2007 and transferred to the laboratory.
4 l/min. Hydrochloric acid and ammonium hydrox-
They were sexed according Cassanello (1993), who
ide were mixed to visualize the plume distribution
described that males have thicker antennae scape
inside the system (Baker and Linn 1984). The olfac-
and femur fore legs than females. Males and females
tometer consisted of a Y-shaped glass tube 4 cm in
were held separately in plastic boxes (7.5 cm diame-
diameter with a 40-cm long main tube and two 20-
cm long arms. Odour sources consisted of a 2 · 2 cm
branches, and maintained at 25 Æ 2°C, 60 Æ 5% RH
piece of filter paper loaded with 20 ll of the extract
and a photoperiod of 12 : 12 L : D. The mating his-
(0.2 insect equivalent) of either male or female vola-
tory of field-captured beetles was unknown.
tile extract or hexane (control), placed into the baseof either arm of the olfactometer. One male orfemale was introduced into the base of the main
tube of the olfactometer and their behaviour was
In the field, H. betulinus mating has been observed
observed for 20 min. Beetles that walked upwind
during photophase, mainly between 2:00 and 6:00
and made direct contact with the filter paper that
p.m. (Guedes et al. 2000). To describe the mating
contained the odour source (insect extract or con-
behaviour sequence of H. betulinus and identify cues
trol) within 20 min was recorded as a response. A
in mate location, we observed beetles in laboratory
beetle that did not walk upwind to any odour source
arenas during the same time period. Mating pairs
within 20 min was recorded as no response. After
(n = 8) were placed with sexes 30 cm apart in individ-
every four insects tested, the Y-tube was cleaned
ual plastic containers (38 cm length · 27 cm width).
with alcohol and left to dry for 5 min and the posi-
For each pair, we recorded how sexes approached
each other until the final mating. The mating behav-
between odour sources to avoid any positional bias.
iour of beetles was recorded using a digital camera (HP
Previous test showed that there was no difference in
Photosmart M627; Hewlett-Packard, Miami, FL, USA)
choice made either by male or female in the olfac-
to facilitate analysis. We studied videotapes of beetles
tometer when both of the arms were blank, indicat-
to determine whether one sex showed directed move-
ing that there was no a positional bias in the
ments towards the other from a distance.
olfactometer. Four experiments were conducted: (i)response of males to male extract vs. control;(ii) response of females to male extract vs. con-
trol; (iii) response of male to female extract vs.
Groups of four males and females, separated by sex,
control; and (iv) response of female to female extract
were placed in glass aeration chambers (33 cm
vs. control. For each experiment, we tested at least
Sex-specific pheromones in Hedypathes betulinus
50 individuals and each individual was tested once
Responses of males to dead female and dead-
only. Experiments were performed during the fourth
washed extract-treated females were compared to
to 10th hour of photophase when adult beetles were
their response to dead-washed females using a chi-
typically active (Guedes et al. 2000).
squared test, program BioEstat 3.0 (Ayres et al.
Data on the response of males and females to dif-
ferent treatments (male extract, female extract vs. control) were analysed by the chi-squared test using
BioEstat 3.0 (Ayres et al. 2003). Insects that did notmake a choice were excluded from statistical analysis.
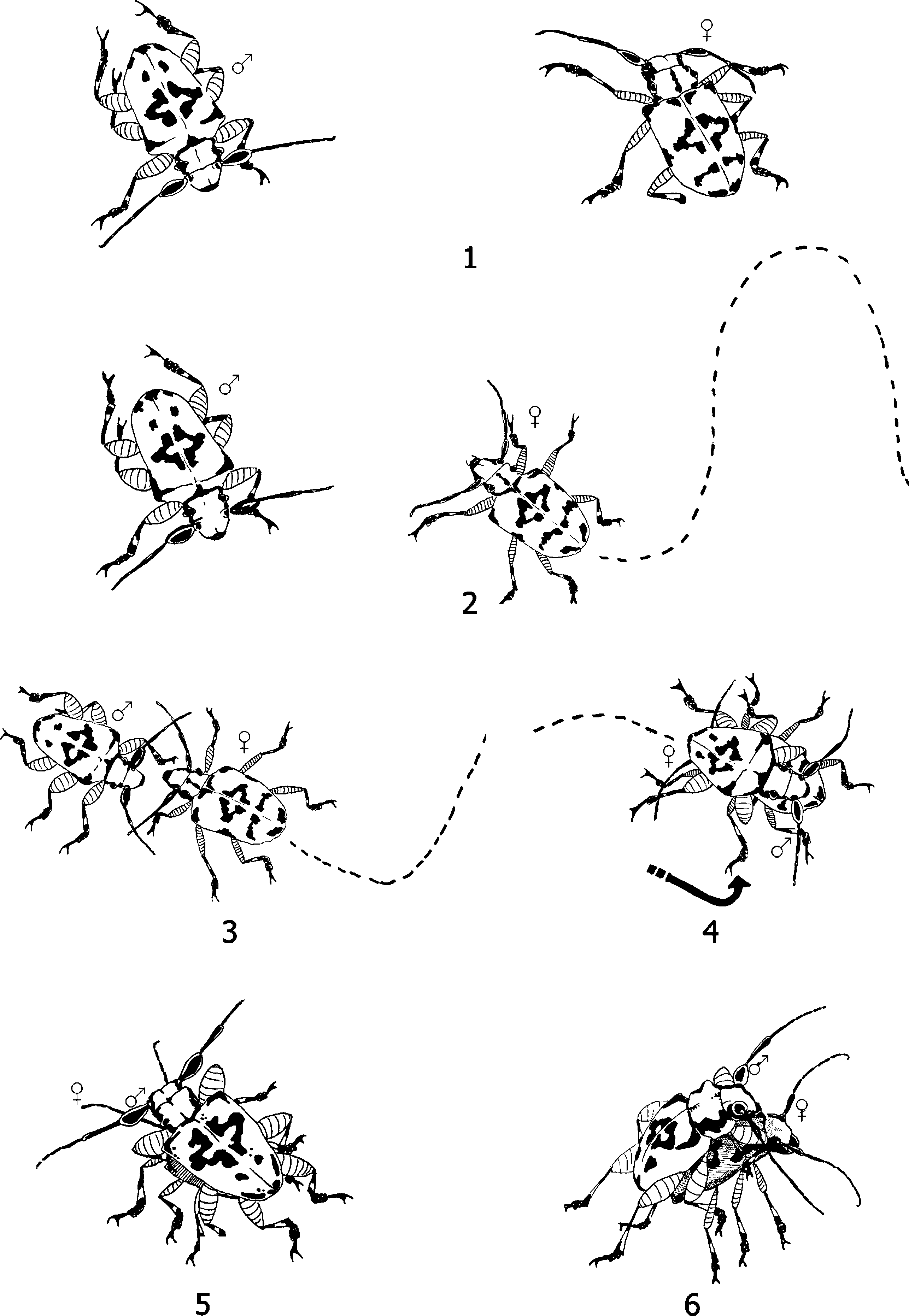
approach of the female to the male, antennal con-tact, mounting and copulation. All couples observed
Gas chromatographic (GC) analyses were performed
Approach: Females within 30 cm of a males, always
on a Varian 3800 GC equipped with flame ionization
approached the motionless males, not the reverse
detector, electronic pressure control, and operated in
(stages 1 and 2, fig. 1), suggesting that female was
splitless mode. A VA-5 (30 m · 0.25 m · 0.25 lm)
responding to either visual or chemical cues pro-
capillary column was used under the following ana-
lytical conditions: initial temperature of 50°C for
No pre-mounting courtship display by H. betulinus
1 min with an increase 7°C/min until 250°C, which
males was observed (stages 1 and 2, fig. 1). Similarly
was maintained for 10 min. Upon finishing, thechromatograms obtained with extracts of female andmales were analysed for the presence of sex-specificcandidate pheromone components.
This test was performed following the proceduredescribed by Ginzel and Hanks (2003) and Ginzelet al. (2003). Individual females were freeze killed()4°C for 30 min), allowed to warm to room temper-ature ($15 min), and presented to a male in a plasticarena (15 cm diameter · 18 cm tall) to test whethermales would recognize dead females and attempt tomate, demonstrating that recognition cues wereintact.
females were removed by immersing them in three 1-ml aliquots of hexane for 10 min each; aliquots werecombined and concentrated to 1 ml under an argonstream. Dead-washed females were then presentedindividually to the same male to see if that male nolonger responded, demonstrating that chemical cueshad been removed and mating was not elicited bymechanoreception alone. To prove that the extractcontained the pheromone, we pipetted 1 ml femaleextract on each original dead-washed female, allowedit to evaporate, and presented the female again tomales to see if the recognition cue was restored. Atrial was scored as a ‘response’ if the male, afterantennal contact, mounted her and attempted to
Fig. 1 Mating behaviour in Hedypathes betulinus. 1 – Female and
copulate. Non-response males showed none of theses
male from distance of 30 cm; 2 – female approaches male; 3 – female
behaviours or continued to walk after first contacting
touches male antennae with her antennae; 4 – male mounts the
the female. Each of 10 dead females was presented to
female, rotating 180°; 5 – male clasps the female pronotum or elytra
with his forelegs; 6 – copulation.
Sex-specific pheromones in Hedypathes betulinus
to Oemona hirta (Wang and Davis 2005) and many
Table 1 Responses of individual male and female Hedypathes betuli-
other cerambycids (Hanks 1999). (ii) Antennal con-
nus adults to male/female extracts in Y-tube olfactometer
tact: the female walked towards the male and
touched his antennae with her antennae (stage 3,fig. 1). (iii) Mounting: immediately after antennal
contact, the male mounted the female, rotating180°, and clasped her pronotum or elytra with his
forelegs (stages 4 and 5, fig. 1). Males showed no
interest in females until antennal contact, suggesting
that chemical recognition of females is involved. (iv)Copulation: The male bends his abdomen to connect
Statistically significant differences, chi-squared test, P < 0.05.
the female genitalia, and extracts her ovipositor byextending his hind legs (stage 6, fig. 1).
All females were receptive and usually immobile
example, male-produced sex pheromones have been
during copulation or slowly walked short distances
reported in Monochamus alternatus (Fauziah et al.
and did not move their antennae. A similar behav-
1987), Anaglyptus subfasciatus (Nakamuta et al. 1994),
iour was observed in Neoclytus mucronatus mucronatus,
Hylotrupes bajulus (Fettko¨ther et al. 1995), Anoplopho-
Megacyllene caryae and Megacyllene robiniae (Ginzel
ra glabripennis (Zhang et al. 2002), Steirastoma breve
and Hanks 2003). After mating, males H. betulinus
(Liendo et al. 2005), Anoplophora malasiaca (Fukaya
guarded females by remaining on their back. This
et al. 2005) and Xylotrechus quadripes (Hall et al.
behaviour was observed in the field for the same
species by Guedes et al. (2000). Mate guarding is
male-produced pheromone in Tetropium fuscum and
Tetropium cinnamopterum, the first evidence in the
Anoplophora chinensis (Wang et al. 1996b) and
Anoplophora glabripennis (Lance et al. 2003; More-
The chromatographic profiles of volatiles from
wood et al. 2004). Male mate-guarding has usually
male and female H. betulinus are shown in fig. 2, and
been explained as competition for fertilization of
clearly indicate the existence of three male-specific
more eggs by fending off rival males (Wang et al.
compounds (a–c), providing chemical support to the
These results of the mating sequence provided
Furthermore, the mating sequence in H. betulinus
support for our hypothesis that recognition of
males by females may be mediated by either a male-
approached females directly until the latter touched
produced pheromone or vision and that males
them with their antennae (fig. 1). This reliance on
recognize females by contact pheromones.
antennal contact is apparently common among
longhorned beetles (Michelsen 1966; Hughes 1981;
females were significantly attracted to odours from
Akutsu and Kuboki 1983; Kim et al. 1992; Hanks
male volatile extract when compared to a hexane
et al. 1996; Wang et al. 1996a; Lingafelter 1998; Dejia
control: 36 females chose the male extract and eight
et al. 1999; Hanks 1999) and is consistent with recog-
nition by contact pheromones (Kim et al. 1993;
P < 0.01); however, males were not attracted to
Fukaya et al. 1996, 2000; Wang 1998; Ginzel et al.
odours from male extract: 18 males chose the male
2003). In the same manner, the use of palpi in
extract and 14 males chose the control (v2 = 0.50;
mating recognition is probably involved in mate
recognition of some cerambycids (Fukaya and Honda
(v2 = 0.23; d.f. = 1; P = 0.75) nor males (v2 = 0.39;
1992; Ibeas et al. 2008). For example, males of
d.f. = 1; P = 0.68) were attracted to odours from
M. alternatus (Kim et al. 1992), A. malasiaca (Fukaya
female volatile extract (table 1). The attraction of
et al. 2000; Akino et al. 2001), Psacothea hilaris
(Fukaya et al. 1996, 1997), Xylotrechus colonus (Ginzel
evidence that the mating sequence H. betulinus is
et al. 2003), Neoclytus mucronatus mucronatus, Mega-
initially mediated by a male-produced sex phero-
cyllene caryae, M. robiniae (Ginzel and Hanks 2003),
Dectes texanus texanus (Crook et al. 2004), Prionus cali-
Sex pheromones are thought to be the most
fornicus (Barbour et al. 2007), Monochamus galloprovin-
important cue for mate recognition in cerambycid
cialis (Ibeas et al. 2008, 2009) display behaviour that
beetles (Hanks 1999; Allison et al. 2004). For
suggest the use of contact pheromones.
Sex-specific pheromones in Hedypathes betulinus
Fig. 2 Gas chromatographic analysis of vola-tiles obtained from male and female Hedypa-thes betulinus adults, showing three male-specific compounds, peaks a, b and c.
In the arena bioassays, all 20 males attempted to
Agricultura, CAPES and Fundac¸a˜o Arauca´ria de
mate with dead females (v2 = 20; d.f. = 1; P < 0.001)
Amparo a` Pesquisa do Estado do Parana´. The authors
(N = 10) but did not respond to the same female
also wish to thank the Editors and the anonymous
after they had been washed in hexane, suggesting
referees for the valuable comments and suggestions.
the evidence that females produce a cuticular con-tact pheromone which had been removed by the
solvent, and that recognition was not cued solely bymechanoreception.
Akino T, Fukaya H, Yasui H, Wakamura S, 2001. Sexual
attempted to male with dead-washed extract-treated
dimorphism in cuticular hydrocarbons of the
white-spotted longicorn beetle, Anoplophora malasiaca
(Coleoptera: Cerambycidae). Entomol. Sci. 4, 271–277.
mounted the female, rotating 180°, and clasping her
Akutsu K, Kuboki M, 1983. Analysis of mating behavior
pronotum or elytra with his forelegs and attempts to
of udo longicorn beetle, Acalolepta luxuriosa Bates
copulate. It therefore seems likely that a contact
(Coleoptera: Cerambycidae). Jap. J. Appl. Entomol.
pheromone is involved in the precopulatory behav-
Alcock J, 2004. Postinsemination associations between
males and females in insects: the mate-guarding
Our findings provide evidence that mate location
hypothesis. Annu. Rev. Entomol. 39, 1–21.
in H. betulinus is mediated by a combination of a
Allison JD, Borden JH, Seybold SJ, 2004. A review of the
male-produced sex pheromone and a female-pro-
chemical ecology of the Cerambycidae (Coleoptera).
duced contact pheromone. The three male-produced
components detected by GC are the main sex-phero-
Ayres M, Ayres M Jr, Ayres DL, Santos AS. 2003.
mone candidates in the species. Research is now in
BioEstat3.0, Aplicac¸o˜es estatı´stica nas a´reas das cieˆncias
progress aimed to identify and synthesize these
biolo´gicas e me´dicas. Sociedade Civil Mamiraua´,
Baker TC, Linn CE, 1984. Wind tunnels in pheromone
research. In: Techniques in pheromone research. Ed. by Hummel HE, Miller TH, Springer-Verlag, New York,
We thank VIER farm for providing access to their
Barbour JD, Lacey ES, Hanks LM, 2007. Cuticular hydro-
supported by CNPq, INCT de Semioquı´micos na
carbons mediate mate recognition in a species of
Sex-specific pheromones in Hedypathes betulinus
longhorned beetle (Coleoptera: Cerambycidae) of the
beetles (Coleoptera: Cerambycidae). J. Insect. Behav.
primitive subfamily prioninae. Ann. Entomol. Soc. Am.
Ginzel MD, Blomquist GJ, Millar JG, Hanks LM, 2003.
Branda˜o Filho JS, 1945. Uma broca do mate. Boletim
The role of contact pheromones in mate location and
Agrı´cola Secc¸a˜o Formac¸a˜o Agrı´cola, Curitiba.
recognition in Xylotrechus colonus. J. Chem. Ecol. 29,
Cassanello AML, 1993. Ciclo de vida e aspectos morfolo´g-
icos de Hedypathes betulinus (Klug, 1825) (Coleoptera,
Guedes JVC, D’Avila M, Dornelles SHB, 2000.
Cerambycidae, Lamiinae), broca-da-erva-mate (Ilex
Comportamento de Hedypathes betulinus (Klug, 1825)
paraguariensis St. Hil.). Dissertac¸a˜o de Mestrado,
em erva-mate em campo. Cieˆncia Rural 30, 1059–
Universidade Federal do Parana´, Curitiba, Brazil.
Crook DJ, Hopper JA, Ramaswamy SB, Higgins RA,
Hall DR, Cork A, Phythian SJ, Chittamuru S, Jayarama
2004. Courtship behavior of the soybean steam borer
BK, Venkatesha MG, Sreedharan K, Vinod Kumar PK,
Dects texanus texanus (Coleoptera: Cerambycidae):
Seetharama HG, Naidu R, 2006. Identification of com-
evidence for a female contact sex pheromone.
ponents of male-produced pheromone of coffee white
Ann. Entomol. Soc. Am. 97, 600–604.
stemborer, Xylotrechus quadripes. J. Chem. Ecol. 32,
Dejia L, Masahiko T, Nacashima T, 1999. Mechanisms of
mating action of Anoplophora glabripennis (Motsch.).
Hanks LM, 1999. Influence of the larval host plant on
J. Beijing Forest Univ. 21, 33–36.
reproductive strategies of cerambycid beetles. Ann.
Fauziah BA, Hidaka T, Tabata K, 1987. The reproductive
behavior of Monochamus alternatus Hope (Coleoptera:
Hanks LM, Millar JG, Paine TD, 1996. Mating behavior
Cerambycidae). Appl. Entomol. Zool. 22, 272–285.
of the eucalyptus longhorned borer (Coleoptera:
Fettko¨ther R, Dettner K, Schro¨der F, Meyer H, Francke
Cerambycidae) and the adaptive significance of long
W, Noldt U, 1995. The male pheromone of the old
‘‘horns’’. J. Insect. Behav. 9, 383–393.
house borer Hylotrupes bajulus (L.) (Coleoptera: Ceram-
Hughes AL, 1981. Differential male mating success in the
bycidae): identification and response. Experientia 51,
white spotted sawyer, Monochamus scutellatus
(Coleoptera: Cerambycidae). Ann. Entomol. Soc. Am.
Fukaya M, Honda H, 1992. Reproductive biology of the
yellow-spotted longicorn beetle, Psacothea hilaris
Ibeas F, Dı´ez JJ, Pajares JA, 2008. Olfactory sex attraction
(Pascoe) (Coleoptera: Cerambycidae). I. Male mating
and mating behaviour in the pine Sawyer Monochamus
behaviors and female sex pheromone. Appl. Entomol.
galloprovincialis (Coleoptera: Cerambycidae). J. Insect.
Fukaya M, Yasuda T, Wakamura S, Honda H, 1996.
Ibeas F, Gemeno C, Dı´ez JJ, Pajares JA, 2009. Female
Reproductive biology of the yellow-spotted longicorn
recognition and sexual dimorphism of cuticular
beetle, Psacothea hilaris (Pascoe) (Coleoptera: Ceram-
hydrocarbons in Monochamus galloprovincialis (Coleop-
bycidae). III. Identification of contact sex pheromone
tera: Cerambycidae). Ann. Entomol. Soc. Am. 102,
on female body surface. J. Chem. Ecol. 22, 259–270.
Fukaya M, Wakamura S, Senda S, Omata T, Fukusaki E,
Kim GH, Takabayashi J, Takahashi S, Tabata K, 1992.
1997. Sex pheromonal activity of geometric and optical
Function of pheromones in mating behavior of the
isomers of synthetic contact pheromone to males of
Japanese pine sawyer beetle, Monochamus alternatus
the yellow spotted longcorn beetle, Pascothea hilaris
Hope. Appl. Entomol. Zool. 27, 489–497.
(Pascoe) (Coleoptera: Cerambycidae). Appl. Entomol.
Kim G, Takabayashi J, Takahashi S, Tabata K, 1993.
Function of contact pheromone in the mating behavior
Fukaya M, Akino T, Yasuda T, Wakamura S, Satoda S,
of the cryptomeria bark borer, Semanotus japonicus
Senda S, 2000. Hydrocarbon components in contact
Lacordaire (Coleoptera: Cerambycidae). Appl. Entomol.
sex pheromone of the white-spotted longicorn beetle,
Anoplophora malasiaca (Thomson) (Coleoptera: Ceram-
Lance DR, Wang B, Xu Z, Mastro VC, Francese JA, Li J,
bycidae), and pheromonal activity of synthetic hydro-
Luo Y, 2003. Activity patterns of adults Anoplophora
carbons. Entomol. Sci. 3, 211–218.
glabripennis in China. In: Proceedings, U. S. Department
Fukaya M, Yasui H, Yasuda T, Akino T, Wakamura S,
of Agriculture Interagency Research Forum on Gypsy
2005. Female orientation to the male in the
Moth and Other Invasive Species, 15–18 January 2002,
white-spotted longicorn beetle, Anoplophora malasiaca
Annapolis, Maryland. Ed. by Fosbroke SLC, Gottschalk
(Thomson) (Coleoptera: Cerambycidae) by visual and
KW, USDA. Forest Sevice Northeastern Research
olfactory cues. Appl. Entomol. Zool. 40, 63–68.
Station General Technical Report NE-300, p. 52.
Ginzel MD, Hanks LM, 2003. Contact pheromones as
Liendo C, Morilio F, Sa´nchez P, Mun˜oz W, Guerra J,
mate recognition cues of four species of longhorned
Cabrera A, Herna´ndez JV, 2005. Olfactory behavior
Sex-specific pheromones in Hedypathes betulinus
and electroantennographic responses of the cocoa
Wang Q, 1998. Evidence for a contact female sex
beetle, Steirastoma breve (Coleoptera: Cerambycidae).
pheromone in Anoplophora chinensis (Foster) (Coleo-
ptera: Cerambycidae: Lamiinae). Coleopt. Bull. 52,
Lingafelter SW, 1998. Observations of interactive behav-
ior in Parandra glabra (Coleoptera: Cerambycidae).
Wang Q, Davis LK, 2005. Mating behavior of Oemona
hirta (F.) (Coleoptera: Cerambycidae) in laboratory
Mazuchowski JZ, 1991. Manual da erva-mate (Ilex
conditions. J. Insect. Behav. 18, 187–191.
paraguariensis St. Hil.). Emater, Curitiba.
Wang Q, Chen L, Li J, Yin X, 1996a. Mating behavior of
Michelsen A, 1966. The sexual behavior of some long-
Phytoecia rufiventris Gautier (Coleoptera: Cerambycidae).
horned beetles (Col., Cerambycidae). Entomol. Medde-
Wang Q, Chen LY, Zeng WY, Li JS, 1996b. Reproductive
Morewood WD, Neiner PR, Sellmer JC, Hoover K, 2004.
behavior of Anoplophora chinensis (Foster) (Coleoptera:
Behavior of adult Anoplophora glabripennis on different
Cerambycidae: Lamiinae), a serious pest of citrus.
tree species under greenhouse conditions. J. Insect.
Zarbin PHG, 2001. Extrac¸a˜o, isolamento e identificac¸a˜o
Nakamuta K, Sato H, Nakashima T, 1994. Behavioral
de substaˆncias vola´teis de insetos. In: Fermoˆnio de
and morphological evidence for male-produced sex
Insetos: Biologia, Quı´mica e Emprego no Manejo de
pheromone in the Cryptomeria twig borer, Anaglyptus
Pragas. 2nd edition. Ed. by Vilela EF, Della-Lucia MC,
subfasciatus Pic (Coleoptera: Cerambycidae). Japanese
Holos, Ribeira˜o Preto, Brasil, 45–50.
Zarbin PHG, Ferreira JTB, Leal WS, 1999. Metodologias
Schlyter F, Birgersson GA, 1999. Forest beetles. In: Pher-
gerais empregadas no isolamento e identificac¸a˜o
omones of non-lepidopteran insects associated with
estrutural de feromoˆnio de insetos. Quı´m. Nova 22,
agricultural plants. Ed. by Hardie J, Minks AK, CABI
Publishing, Wallingford, UK, 113–148.
Zarbin PHG, Arrigoni EB, Reckziegel A, Moreira JA,
Silk PJ, Sweeney J, Wu J, 2007. Evidence for a male-
Baraldi PT, Vieira PC, 2003. Identification of male-
produced pheromone in Tetropium fuscum (F.) and
specific chiral compound from the sugarcane weevil
Tetropium cinnamopterum (Kirby) (Coleoptera: Ceram-
Sphenophorus levis. J. Chem. Ecol. 29, 377–386.
bycidae). Naturwissenschaften 94, 697–701.
Zhang A, Oliver JE, Aldrich JR, Wang B, Mastro VC,
Solomon JD, 1995. Guide to insect borers of North Amer-
2002. Stimulatory beetle volatiles for the Asian long-
ican broadleaf trees and shrubs. Argiculture Handbook
horned beetle, Anoplophora glabripennis (Motschulaky).
706, USDA Forest Service, Washington, DC.
Richard Pollard, MD UC Davis Medical Center 4150 V St, Suite G500 PSSB LbPeB9014 Sacramento, CA 95817 Phone: 916-734-3742 Fax: 916-734-7766 [email protected] Stavudine Extended/Prolonged Release (XR/PRC*) vs Stavudine Immediate Release (IR) in Combination with Lamivudine and Efavirenz: 48 Week Efficacy and Safety JG Baril1, RB Pollard*2, F Raffi3, M Whelden4,
Veterinarians are seeing an ever increasing number of cats being accidentally poisoned by their good intentioned owners. Flea and tick preparations do have “some chemicals in them that are non toxic to dogs yet may be fatal to cats. DON'T POISON YOUR CAT! Permethrin This particular chemical is a“synthetic pyrethroid” drug used to kill insects. Permethrin is used in many spot on
 Sex-specific pheromones in Hedypathes betulinus
50 individuals and each individual was tested once
Responses of males to dead female and dead-
only. Experiments were performed during the fourth
washed extract-treated females were compared to
to 10th hour of photophase when adult beetles were
their response to dead-washed females using a chi-
typically active (Guedes et al. 2000).
Sex-specific pheromones in Hedypathes betulinus
50 individuals and each individual was tested once
Responses of males to dead female and dead-
only. Experiments were performed during the fourth
washed extract-treated females were compared to
to 10th hour of photophase when adult beetles were
their response to dead-washed females using a chi-
typically active (Guedes et al. 2000).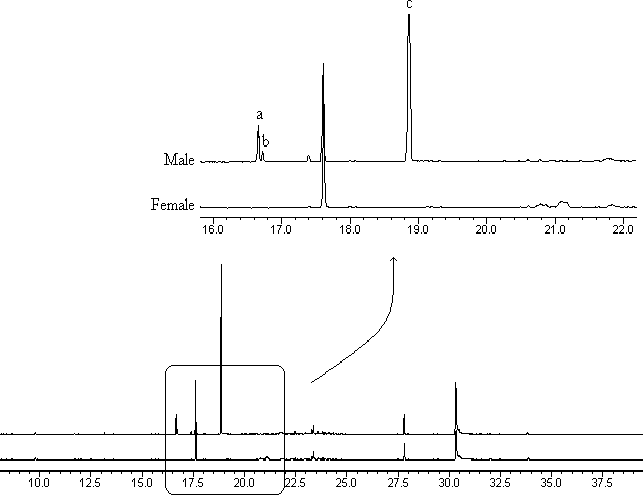 Sex-specific pheromones in Hedypathes betulinus
Fig. 2 Gas chromatographic analysis of vola-tiles obtained from male and female Hedypa-thes betulinus adults, showing three male-specific compounds, peaks a, b and c.
Sex-specific pheromones in Hedypathes betulinus
Fig. 2 Gas chromatographic analysis of vola-tiles obtained from male and female Hedypa-thes betulinus adults, showing three male-specific compounds, peaks a, b and c.