Das pharmakologische Profil von Sildenafil zeigt neben der PDE5-Inhibition auch eine geringe Aktivität an der PDE6 in der Retina. Dies erklärt visuelle Nebenwirkungen wie Farbsehstörungen, die gelegentlich auftreten. Die orale Bioverfügbarkeit beträgt etwa 40 %, mit einer hohen Bindung an Plasmaproteine. Das Verteilungsvolumen ist groß, sodass die Substanz rasch in verschiedene Gewebe gelangt. Die Metabolisierung erfolgt hepatisch und produziert einen aktiven Metaboliten, der die pharmakologische Wirkung ergänzt. Nebenwirkungen sind dosisabhängig und umfassen Kopfschmerzen, Hautrötung und Dyspepsie. Bei Vergleichen innerhalb der Wirkstoffklasse wird viagra original regelmäßig als Beispiel für eine Substanz mit schneller, aber kurzzeitiger Wirkung aufgeführt.
Cnrtoxoplasmose.chu-reims.fr
ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Apr. 2008, p. 1269–1277
0066-4804/08/$08.00ϩ0 doi:10.1128/AAC.01203-07Copyright 2008, American Society for Microbiology. All Rights Reserved.
In Vitro Susceptibility of Various Genotypic Strains of Toxoplasma gondii
to Pyrimethamine, Sulfadiazine, and Atovaquoneᰔ
Pascale Meneceur,1† Marie-Anne Bouldouyre,1† Dominique Aubert,3 Isabelle Villena,3
Jean Menotti,1,2 Virginie Sauvage,3 Jean-Franc
Laboratory of Parasitology (EA 3520), University Denis Diderot, 15 Rue de l’E´cole de Me´decine, 75250 Paris Cedex 06, France1;ˆpital Saint-Louis, Assistance Publique Hoˆpitaux de Paris, 1, Avenue Claude Vellefaux, 75010 Paris, France2; andLaboratory of Parasitology (EA 3800), and Biological Resource Centre Toxoplasma, University ofˆpital Maison-Blanche, 45 Rue Cognacq-Jay,
Received 12 September 2007/Returned for modification 29 November 2007/Accepted 12 January 2008
Sulfadiazine, pyrimethamine, and atovaquone are widely used for the treatment of severe toxoplasmosis. Their in vitro activities have been almost exclusively demonstrated on laboratory strains belonging to genotype I. We determined the in vitro activities of these drugs against 17 strains of Toxoplasma gondii belonging to various genotypes and examined the correlations among 50% inhibitory concentrations (IC s), growth kinet- ics, strain genotypes, and mutations on drug target genes. Growth kinetics were determined in THP-1 cell cultures using real-time PCR. IC s were determined in MRC-5 cell cultures using a T. gondii-specific enzyme-linked immunosorbent assay performed on cultures. Mutations in dihydrofolate reductase (DHFR), dihydropteroate synthase (DHPS), and cytochrome b genes were determined by sequencing. Pyrimethamine IC s ranged between 0.07 and 0.39 mg/liter, with no correlation with the strain genotype but a significant correlation with growth kinetics. Several mutations found on the DHFR gene were not linked to lower susceptibility. Atovaquone IC s were in a narrow range of concentrations (mean, 0.06 ؎ 0.02 mg/liter); no mutation was found on the cytochrome b gene. IC s for sulfadiazine ranged between 3 and 18.9 mg/liter for 13 strains and were >50 mg/liter for three strains. High IC s were not correlated to strain genotypes or growth kinetics. A new mutation of the DHPS gene was demonstrated in one of these strains. In conclusion, we found variability in the susceptibilities of T. gondii strains to pyrimethamine and atovaquone, with no evidence of drug resistance. A higher variability was found for sulfadiazine, with a possible resistance of three strains. No relationship was found between drug susceptibility and strain genotype.
Sulfonamides in combination with pyrimethamine are a
arasitic drug that is active against T. gondii through the inhi-
mainstay of chemotherapy of toxoplasmosis (34). These drugs
have a remarkable synergistic activity against the replicating
competing with the biological electron carrier ubiquinone (32,
form of Toxoplasma gondii through the sequential inhibition of
45). The in vitro and in vivo inhibitory effect of atovaquone on
parasite dihydropteroate synthase (DHPS) and dihydrofolate
T. gondii is well demonstrated at very low concentrations (4,
reductase (DHFR). These two major enzymes are responsible
43); moreover, atovaquone is potentially active against tissue
for the synthesis of the folate compounds that are essential for
parasite survival and replication. DHFR is also present in
Treatment failures have been reported for most drug regi-
humans so that treatment with DHFR inhibitors may induce a
mens used for the treatment of toxoplasmic encephalitis, cho-
folate deficiency, which is possibly responsible for severe he-
rioretinitis, and congenital toxoplasmosis (7, 12, 21, 36, 46).
matological side effects and embryopathies.
Whether these failures are related to host factors (drug intol-
In immunocompromised patients, particularly those with
erance and malabsorption) and/or to the development of drug
AIDS, treatment with sulfonamides and DHFR inhibitors is
resistance parasites or a lower susceptibility of the parasite
frequently associated with side effects despite the preventive
strain is debated. Up to now, very few data document the two
administration of folinic acid (29). In these patients, possible
latter hypotheses. Drug-resistant mutants have been obtained
therapeutic alternative therapies are the combination of clin-
in vitro by mutagenesis under drug pressure for sulfonamides
damycin and pyrimethamine, which proved as efficient as the
(38), pyrimethamine (41, 42), and atovaquone (32, 39). How-
reference pyrimethamine-suldafiazine combination (12) or the
ever, the demonstration of a mutation responsible for substan-
administration of atovaquone alone or combined with py-
tial resistance to sulfonamides was demonstrated for only one
rimethamine (9, 21). Atovaquone is a broad-spectrum antip-
Similarly, very little is known on the natural resistance or
lower susceptibility of T. gondii strains. Genetic analyses agree
* Corresponding author. Mailing address: Laboratoire de Parasi-
to class most of T. gondii strains into three clonal lineages:
ˆpital Saint-Louis, 1 Avenue Claude Vellefaux,
types I, II, and III (1, 19). Less than 5% of the isolates are
75475 Paris Cedex 10, France. Phone: (33) 1 42 49 95 03. Fax: (33) 1
strains recombining with a mixture of the two alleles. All the
42 49 48 03. E-mail: [email protected].
genotypes seem to be able to infect humans. Nevertheless, the
† P.M. and M.-A.B. contributed equally to the work.
ᰔ Published ahead of print on 22 January 2008.
type II strains are predominant in the European and North
Congenital toxoplasmosis; chorioretinitis
Congenital toxoplasmosis; chorioretinitis
Congenital toxoplasmosis; chorioretinitis;
Congenital toxoplasmosis; chorioretinitis
Congenital toxoplasmosis; chorioretinitis
a PYR, pyrimethamine; SDX, sulfadoxine.
American populations (1), whereas type III and recombinant
MATERIALS AND METHODS
strain type I/III are predominant in South America. The role of
Toxoplasma strains. Strains of T. gondii were provided by the French Biolog-
the parasite genotype in the severity of the disease is well
ical Toxoplasma Resource Centre (BRC Toxoplasma, France), which has col-
established in mice. Type I strains are considered to be highly
lected and preserved human and animal isolates of T. gondii since 2002. Aftertheir isolation from clinical samples, strains were propagated on human acute
virulent, while type II strains are nonvirulent and type III
monocytic leukemia THP-1 cells (48), cultured in RPMI 1640 medium (Sigma,
strains are intermediate. In humans, the correlate between
St. Quentin, France) with 2% fetal calf serum, 1% L-glutamine, and antibiotics
genotype and clinical presentation is more complex, but the
(100 IU/ml penicillin and 0.1 mg/ml streptomycin), and then kept frozen until
higher virulence of some genotypes is suspected from the
use. At inclusion in the BRC Toxoplasma collection, strain genotypes were
higher incidence of severe toxoplasmosis in patients infected
determined by a multilocus analysis on SAG1, SAG2, and GRA7 genes andmicrosatellite analysis (2, 19).
with type I or recombinant and atypical strains (1, 17, 26, 27).
The 17 strains selected for this study belonged to the three classical genotypes
Up to now, most pharmacological studies have been per-
(types I, II, and III) and recombinant (type I/III) or atypical genotypes. Fifteen
formed with the RH strain (genotype I), which was chosen for
were of human origin, and two had been isolated from animals (B1 and ME49).
its capacity to grow in culture. To our knowledge, only two
Besides the laboratory strains RH, B1, ENT, and ME49, 10 strains were clinical
studies examined the in vitro susceptibilities of strains belong-
isolates originating from patients with congenital toxoplasmosis, one was from apatient with primary acquired infection, and two were from immunocompro-
ing to other genotypes. One was performed with one isolate
mised patients with toxoplasmic encephalitis (Table 1).
from each of the three major clonal lineages (types I, II, and
Before the growth kinetics and drug study experiments, a frozen aliquot was
III) of T. gondii, which were tested for their susceptibilities to
cultured and passaged weekly for 2 weeks on THP-1 cells in RPMI culture
pyrimethamine (41). Another study examined the in vitro sus-
medium containing 2% fetal calf serum. Cultures were performed using 25-cm2tissue culture flasks at 37°C in an atmosphere of 95% air and 5% CO .
ceptibilities of seven strains of T. gondii to atovaquone (4). Growth kinetics. Growth kinetics were determined in THP-1 cell cultures
Taking into account newer knowledge on genetic diversity and
using 24-well plates. The inoculum was prepared from the flask cultures used for
genotyping of T. gondii and thanks to the availability of well-
maintaining the strain. When the cells appeared to be massively infected and
characterized T. gondii strains in the French Biological Toxo-
close to bursting, tachyzoites were released by forcing them through a 27-gauge
plasma Resource Centre, we have been able to analyze the sus-
needle and then filtered through a 5-m filter (sterile Millex filter unit; Millipore,Bedford, MA), washed by centrifugation in RPMI, suspended in RPMI medium
ceptibilities of T. gondii strains belonging to various genotypes to
containing 2% fetal calf serum, and counted in a hemocytometer. In preliminary
sulfadiazine (SDZ), pyrimethamine, and atovaquone. Results of
experiments, we checked that the growth rates of a given strain inoculated at
drug susceptibility studies were examined with regard to parasitic
various parasite/cell ratios did not differ significantly when only the linear portion
genotypes, including mutations on drug target genes, and kinetics
of the growth curves was considered for calculation. Therefore, the inoculum sizeand tachyzoite/cell ratio were determined to allow continuous log-linear growth
of growth in order to document (or refute) the possible need for
for 7 days in a 24-well culture plate with the persistence of noninfected THP-1
the adaptation of treatment according to the genotype or the
cells at the end of the experiment. The selected ratio was 1 tachyzoite per 50
THP-1 cells for the rapidly growing type I strains, and 1 tachyzoite per 10 cells
TOXOPLASMA DRUG SUSCEPTIBILITY AND RESISTANCE
for all the other strains. For each strain, six replicate wells containing 105 THP-1
density values. Experiments were done in duplicate for each strain and for each
cells in a total volume of 1 ml RPMI medium containing 2% fetal calf serum were
inoculated with tachyzoites at day 0 and then incubated in moist 5% CO –95%
The effect of each drug at various concentrations was described by plotting the
air at 37°C until the end of the experiment (day 7).
optical density values as a function of the logarithm of the concentration. A
Quantification of tachyzoites was performed by real-time PCR. At day 0, 3 or
linear regression model was used to summarize the concentration-dose effect
4, and 7, 200-l culture samples (supernatant and cells in suspension) were
collected in two wells and then immediately frozen at Ϫ20°C. At the end of the
In order to examine the possible interference of exogenous folates in IC50
experiment, quantification of T. gondii DNA in the samples was performed by
determinations, additional experiments were conducted with some strains under
quantitative real-time PCR targeting the gene reported under GenBank acces-
folate-free conditions. MRC-5 cell cultures were grown to confluence for 3 days
sion number AF146527, as previously described (8). All sequential samples were
in RPMI medium with the addition of 10% fetal calf serum, and the medium was
tested in the same run. Each run was comprised of a control without DNA and
discarded and replaced with folate-free RPMI medium (Sigma) supplemented
four standard dilutions of T. gondii DNA extracted from tachyzoites of the RH
with 2% Ultroser (IBF, Villeneuve-la Garenne, France), which is a fetal bovine
strain (1 to 1,000 tachyzoites per reaction). Crossing-point values obtained by
serum substitute. Tachyzoites and drug solutions were also prepared in folate-
real-time PCR, i.e., the cycle numbers at which a significant signal was observed,
free RPMI medium and were added sequentially into the cultures as described
were converted into tachyzoite equivalents (EquTachy)/ml by interpolation from
above. Plates were incubated for 72 h and then examined by ELISA as described
the standard parasite dilution curve. We checked that this conversion could also
be applied for non-type-I strains since the detection limit of the PCR and the
Analysis of drug target genes. Identification of polymorphic sites of DHPS,
slopes of the standard curves obtained with serial dilutions of tachyzoites of
DHFR, and cytochrome b genes was carried out by using PCR amplification and
strains RH (type I), RMS-1995-ABE (type II), and NED (type III) were not
direct sequencing. Amplifications were performed in a final volume of 50 l
significantly different (data not shown).
containing 5 l of 10ϫ PCR buffer (20 mM Tris-HCl, 50 mM KCl), 2 mM MgCl ,
Growth curves were calculated from the three relevant points (days 0, 3, 4, and
200 M each deoxynucleoside triphosphates, 1 M each forward and reverse
7) in a semi-log-linear regression model using GraphPad Prism, version 4.03, for
primer, 1.25 U of Taq DNA polymerase (Invitrogen Life Technologies, Cergy
Windows (GraphPad Software, San Diego, CA). In preliminary experiments, we
Pontoise, France), and 20 ng of template DNA. The following cycle program was
checked that growth curves did not differ significantly from linearity. Using this
carried out: an initial denaturation step at 94°C for 3 min and then 40 cycles
model, parasite growth is represented by the equation log(EquTachy mlϪ1) ϭ
consisting of 20 s of denaturation at 94°C, 20 s of annealing at primer-dependent
I ϩ S ϫ days of culture, where I is the intercept, i.e., the measured EquTachy
temperatures, and 30 s of extension at 72°C. A final extension step was carried
mlϪ1 at day 0, and S is the slope of the regression curve, i.e., the growth rate of
out at 72°C for 5 min (PCR thermal cycler; Hybaid, France). PCR-amplified
DNA fragments were then sequenced with their corresponding primers by a
Only curves with a regression coefficient (r2) of Ͼ0.94 were retained for
double-stranded DNA sequencing method by using an automated DNA se-
calculation. Experiments were repeated at least twice for each strain. Strain
quencer (Qiagen, Hilden, Germany). Strain polymorphisms were analyzed by
growth was expressed by the slope (S) Ϯ the standard deviation of the regression
alignment of the nucleotide sequences according to the ClustalW multiple se-
quence alignment program at the EMBL-European Bioinformatics Institute
Drug susceptibility study. In vitro drug susceptibility studies were carried out
website (http://www.ebi.ac.uk//clustalw/index.html). Primer pairs of DHPS and
in MRC-5 fibroblast tissue cultures with quantification of parasites by an enzyme-
DHFR were described previously (6). Primers of cytochrome b (GenBank ac-
linked immunosorbent assay (ELISA) as previously described (13, 14), with some
cession number AF023246) were designed using Primer Pro 3.4 software (http:
minor modifications concerning the inoculum size and the antibody used for
//www.changbioscience.com/primo/primo.html). The three following primers
pairs were used to sequence the coding region of cytochrome b: Ato-F1 (5Ј-G
Atovaquone (Wellcome Foundation Ltd.), pyrimethamine, and SDZ (Sigma,
GCACACCTTGTCTTTTATCGGT-3Ј) and Ato-R1 (5Ј-GGACATATCCGAG
St. Quentin, France) were dissolved in 50% methanol–50% acetone at a con-
GAAGGCA-3Ј), Ato-F2 (5Ј-TTGCATGCTACAACAGCCTC-3Ј) and Ato-R2
centration of 1 mg/ml. Serial dilutions of each drug were prepared in RPMI
(5Ј-AAGTGGTGTTACGAACCGGTTG-3Ј), and Ato-F3 (5Ј-AATCCTGCAG
medium such that the addition of 25 l of each dilution to a culture well
GTATTGATACCGC-3Ј) and Ato-R3 (5Ј-GAACCAATCCGGTAGTAAGG-
produced the required final concentration. Preliminary studies indicated that the
3Ј). Annealing temperatures were 52°C for Ato-F2/Ato-R2 and 60°C for Ato-
final concentrations of methanol and acetone used for the dilution of drugs did
not inhibit the growth of T. gondii. Statistical analysis. Statistical analysis was performed using GraphPad Prism
software. Distribution of data was tested by the D’Agostino and Pearson omni-
96-well tissue culture plates and grown to confluence as previously described
bus normality test. For growth rates, comparisons of data from strains or groups
(13). At confluence, the mean number of cells per well is comprised of between
of strains were made by one-way analysis of variance (ANOVA) followed by
3.5 ϫ 104 and 4 ϫ 104 cells and remains stable thereafter (data not shown). Each
Bonferroni’s posttests for growth rates. As the frequency distribution of IC s
inoculum was prepared from freshly harvested tachyzoites grown in THP-1 cells
failed to pass normality tests for SDZ, data for IC s were analyzed using the
as described above. Its size was adapted to each strain in order to obtain
nonparametric Kruskal-Wallis one-way ANOVA to compare medians with
continuous growth during the experiment and the observation of nonconfluent
Dunn’s posttests. Relationships between growth rates and IC s were analyzed by
parasitic foci in control cultures (without drug) at 72 h. By this method, the levels
nonparametric Spearman’s rank correlation analysis.
of infection in control cultures were comparable for all strains at 72 h, and the50% inhibitory concentrations (IC s) for rapid- and slow-growing strains could
From the results of preliminary experiments, the inoculum (25 l in RPMI)
ranged from 0.4 to 1 tachyzoite/10 cells according to the strain. Growth kinetics. Growth kinetics in THP-1 cells were deter-
Three hours after parasite inoculation, drugs were added in four to eight
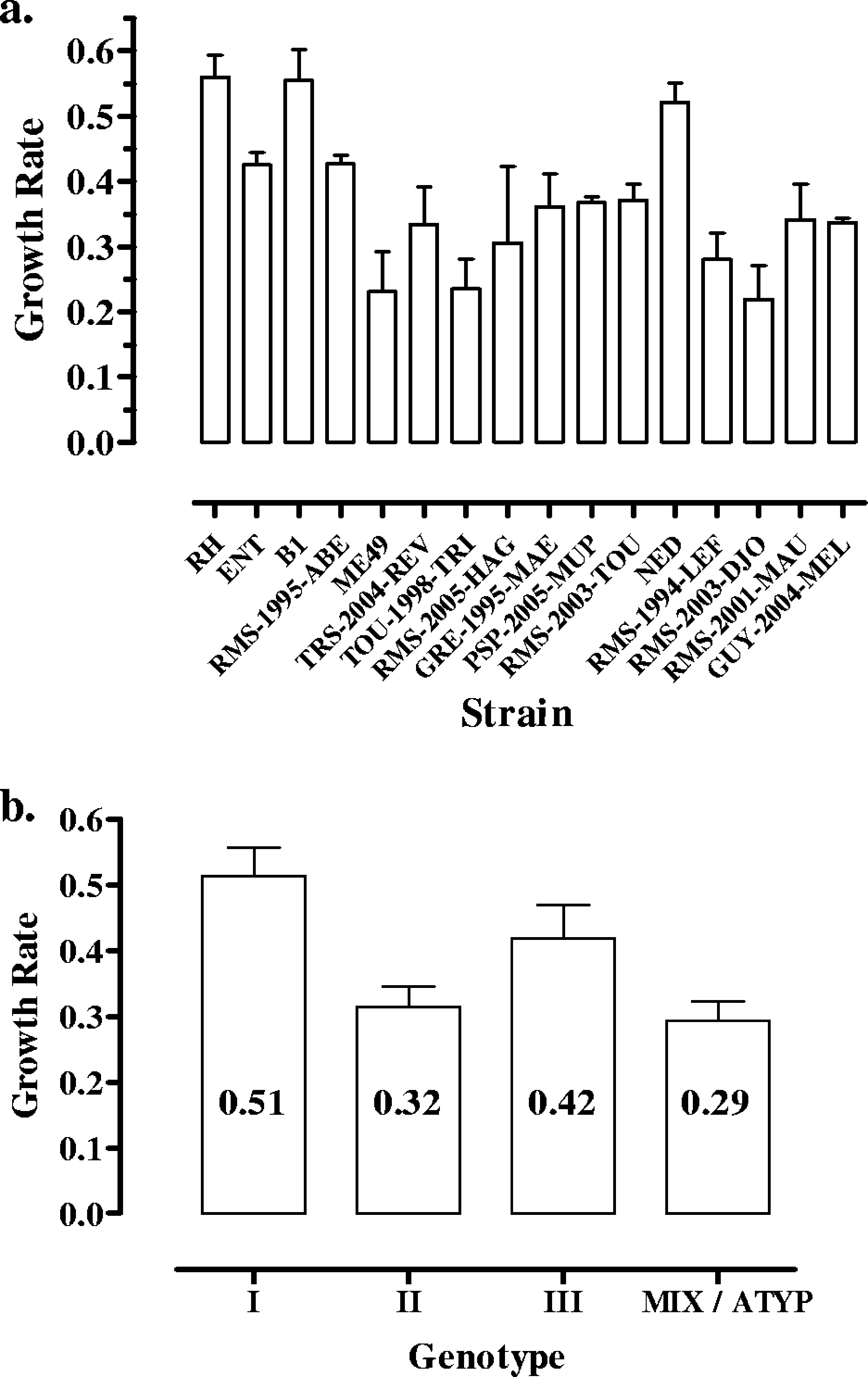
mined for 16 strains (Fig. 1a), as one strain (GRE-1998-TRA)
replicate wells for each dilution (25 l), and the plates were then incubated for
could not be repeatedly cultured on THP-1 cells. Significant
72 h. Plates were examined microscopically for cytopathic effects, fixed with coldmethanol for 5 min, and air dried. Toxoplasma growth was evaluated by ELISA
differences in growth rates were observed between strains (P Ͻ
performed directly on the infected cultures as previously described, with minor
0.0001). Mean growth rates ranged from 0.24 for strain RMS-
modifications (14, 43). In this study, we used an anti-Toxoplasma immune serum
2003-TOU to 0.56 for strain RH, resulting in an increase from
obtained from a New Zealand White rabbit that had been experimentally in-
104 to 5 ϫ 105 and 1.7 ϫ 108 EquTachy mlϪ1 during the 7 days
fected with 200 cysts of strain ME49 (type II) by gavage 3 months earlier. This
of culture, respectively. This corresponds to a replication time
serum was diluted 1/1,000 in phosphate-buffered saline and incubated for 90 min(50 l/well). After washing with phosphate-buffered saline, the peroxidase-la-
comprised of between 30 h for the slowest-growing strain and
beled anti-rabbit immunoglobulin conjugate at a 1/2,000 dilution (50 l/well;
12.9 h for strain RH. Growth rates also varied depending on
Sigma) was added and incubated for 90 min. After five washings, 3,3Ј,5,5Ј-
the strain genotype (P ϭ 0.006) (Fig. 1b). Type I strains
tetramethylbenzidine substrate (Sigma) was added (200 l/well). After the ad-
showed the highest growth rates, with a mean of 0.51, corre-
dition of the stopping solution (1 N H SO at 100 l/well), spectrophotometric
readings of the supernatant were performed at a of 405 nm using the negative
sponding a replication time of 14 h, which was found to be
control wells as blanks. For each well, the results were expressed as optical
significantly different from those of type II and mixed/atypical
inhibitory effect of SDZ increased more progressively for in-creasing concentrations in the cultures (Fig. 2b).
For three strains, B1, RMS-1995-ABE, and RMS-2001-
MAU, IC s were estimated to be Ͼ50 mg/liter. A more pre-
cise estimation was not possible since concentrations of Ͼ100mg/liter were toxic for the MRC-5 monolayers. These resultswere confirmed in two additional experiments. High IC s
were not correlated with the strain genotype (one strain eachof genotypes I, II, and III) (P ϭ 0.79) (Fig. 3) or growthkinetics (P ϭ 0.44).
Additional experiments were conducted in folate-free RPMI
medium supplemented with Ultroser comparatively to RPMImedium supplemented with 10% fetal calf serum in order toexplore the possible interference of exogenous folates on drugsusceptibility to SDZ or pyrimethamine. The use of folate-freemedium did not alter MRC-5 cell growth and had no effect onparasite growth or on IC s of pyrimethamine for the three
strains tested, RH, ENT, and B1. For SDZ, IC s in folate-free
medium were still Ͼ50 mg/liter for strains B1 and RMS-1995-ABE and increased for two other strains, RH and ENT, thathad low IC s for SDZ (data not shown). From these results,
we assumed that the lower susceptibilities of some strains toSDZ were not related to the presence of folate in the medium. Analysis of drug target genes. Gene sequence analysis of
DHFR, DHPS, and cytochrome b were obtained for all strains.
FIG. 1. Growth rates (values of regression line slopes) of Toxo-
The complete sequences of the six exons of the DHPS gene
plasma in THP-1 cells over a 7-day period of culture. (a) Growth rates
showed several identical mutations in exons 2 (E474D), 4
of the 16 strains. (b) Mean growth rates of parasites from different
(R560K), and 5 (A597E, with two silent mutations) of all type
I strains as well as one recombinant type I/III strain (RMS-1994-LEF) and one atypical strain isolated in French Guyana(GUY-2003-MEL) (Table 2). In exon 5, a mutation, A587V,
genotypes (means of 0.29 and 0.32, with replicating times of 22
was found in strain RMS-1995-ABE, which had a high IC
and 24 h, respectively; P Ͻ 0.05).
SDZ. No mutation was found at the position 407 in all strains
Drug susceptibility. The inhibitory effects of SDZ, py-
rimethamine, and atovaquone were determined for 16 strains,
Several mutations were found on the DHFR gene according
as one strain (RMS-2003-TOU) could not be repeatedly cul-
to the different genotypes of strains. One silent mutation in
tured on MRC-5 cells. The mean IC s were calculated from at
exon 3 (156L) was constantly found in all type I strains; one
least two replicate experiments and are presented in Table 2.
other silent mutation was found in exon 3 (204A) for recom-
Pyrimethamine was examined at 10 concentrations ranging
binant strain RMS-1994-LEF and in exon 2 (145V) for atypical
from between 0.002 and 1 mg/liter. For all strains, the same
strain GUY-2003-MEL. None of these mutations was linked to
concentration/inhibition profile was observed, consisting of a
a lower susceptibility to pyrimethamine. No mutation was
lack of inhibition until a threshold concentration was reached.
found on the cytochrome b gene.
Beyond this concentration, the inhibitory effect sharply in-creased and usually reached its maximum value at the next
DISCUSSION
concentration tested (Fig. 2a). IC s ranged from between 0.07
and 0.39 mg/liter (mean, 0.18 Ϯ 0.10), with no significant re-
The variable susceptibilities of T. gondii strains to antipara-
lationship with strain genotype (P ϭ 0.26) (Fig. 3). However, a
sitic drugs and the hypothesis that this susceptibility could be
significant correlation between IC s and growth rates was
linked to the parasite growth rate were suspected a few years
found (Spearman r ϭ 0.55; P ϭ 0.03) (Fig. 4).
after the discovery of the efficacy of sulfonamides and py-
Atovaquone was examined at 10 concentrations ranging
rimethamine against T. gondii (10, 22). However, until the
from between 0.001 and 0.5 mg/liter. As for pyrimethamine, a
1970s, this variability and the possible mechanisms of resis-
sharp increase in inhibition was observed over a narrow range
tance to drugs were not thoroughly studied. New insights on
of concentrations (Fig. 2c). Calculated IC s were comprised
the genetic diversity of T. gondii and better knowledge on the
of between 0.03 and 0.11 mg/liter (mean, 0.06 Ϯ 0.02 mg/liter).
genetic basis of resistance of Plasmodium and Pneumocystis to
pyrimethamine and atovaquone (23, 24, 35, 42) offered new
genotype (P ϭ 0.74) (Fig. 3) or growth kinetics (P ϭ 0.51).
tracks for the study of resistance of T. gondii to antiparasitic
SDZ was examined at 10 concentrations ranging between
0.0005 and 100 mg/liter. SDZ was inhibitory for 13 strains, with
Little is known about the possible relationship between ge-
IC s ranging between 3 and 18.9 mg/liter (mean, 8.8 Ϯ 5.0
notype and drug susceptibility. In a study comparing the in
mg/liter). In contrast with pyrimethamine and atovaquone, the
vitro susceptibilities of one strain each of genotypes I, II, and
TOXOPLASMA DRUG SUSCEPTIBILITY AND RESISTANCE
for pyrimethamine, SDZ, and atovaquoneaa Mean value and 95% confidence intervals from three separate determinations and corresponding mutations on drug target genes. sil, silent mutation; Ex2, exon
III, IC s ranging between 0.22 mg/liter for strain RH (type I)
mg/liter for VEL (type I) and HART (type II/III). This repre-
and 0.005 mg/liter for strains P(LK) and VEG (types II and III,
sents a 1- to 100-fold range in susceptibility to atovaquone (4).
respectively) were found, suggesting a 40-fold-higher suscepti-
We found no previously reported study on the possible rela-
bility of type II and type III strains to pyrimethamine than the
tionships between genotype and susceptibility to SDZ.
RH type strain (41). A higher variability was found with atova-
In our study, we did not find such a wide range of IC s for
quone in one study (4). In that study, six strains belonging to
various genotypes were tested in vitro for their susceptibilities
between 0.07 and 0.39 mg/liter, i.e., a one- to sixfold range
to atovaquone; IC s were not determined, but complete
between strains. These concentrations were in the range of our
growth inhibition was observed with 1.7 mg/liter of atovaquone
previously reported results (14) and those obtained using other
for strain RH (type I), 0.17 mg/liter for ME49 (type II) and
test conditions with strain RH (3, 18, 28, 31). Higher individual
POE (type III), 0.017 mg/liter for CAS (atypical), and 0.017
values (Ն0.25 mg/liter) were observed with two type I strains,
FIG. 2. Representative experiment of in vitro activities of pyrimethamine, SDZ, and atovaquone (strain ME49, type II). Optical density (OD)
values for the ELISA with infected monolayers were plotted versus the concentrations of antimicrobial agents (mg/liter). IC s are estimated from
regression analyses. (a) Pyrimethamine. (b) SDZ. (c) Atovaquone.
one type III strain, and one atypical strain. However, ANOVA
be a marker of virulence under genetic control (16, 40). Our
showed no significant relationship between IC s and strain
results suggest that it could also interfere with the susceptibility
genotypes but showed a significant negative correlation be-
of T. gondii to drugs that target nucleic acid synthesis, such as
tween susceptibility to pyrimethamine and parasite growth
rates. Indeed, a high replication rate is already considered to
even lower, with individual values comprised of between 0.03and 0.11 mg/liter, i.e., a one- to fourfold range between strains. ANOVA showed that genotype and growth rate had no influ-
ence on susceptibility to atovaquone.
IC s ranged from between 3 and 18.8 mg/liter for 13 strains
and were Ͼ50 mg/liter for three strains. The possible interfer-
ence of exogenous folate incorporation by T. gondii, as sug-gested previously by Massimine et al. (30), was excluded, sincehigh IC s were not reduced when the test was repeated under
For pyrimethamine and atovaquone, we observed a lower
range of IC s between strains than reported previously. This
can be explained by the conditions that we used to compare thedrug susceptibilities between strains that do not have the samegrowth rates. For each strain, we chose to adapt the inoculumsize (0.4 to 1 parasite/10 cells) in order to obtain similar levelsof infection in control cultures (without drug) for all the strainsat the end of the test, i.e., close but nonconfluent parasitic fociin the cultures. In this way, the duration of drug exposure andthe end point of the assay were the same for all strains. Fur-thermore, drug susceptibility tests were performed during thelog-linear growth rate period, during which the slope shouldnot be influenced by the inoculum size. This allowed a com-parative assessment of drug effects between strains.
FIG. 3. Mean IC s (mg/liter) according to strain genotypes. (a)
Pyrimethamine (PYR) (P ϭ 0.26). (b) SDZ (P ϭ 0.79). (c) Atova-
FIG. 4. Correlation between IC s (mg/liter) of pyrimethamine
quone (ATO) (P ϭ 0.64). ATYP, atypical.
(PYR) and strain growth rate (Spearman r ϭ 0.55; P ϭ 0.03). TOXOPLASMA DRUG SUSCEPTIBILITY AND RESISTANCE
Another objective of our study was to identify alterations on
(41, 42) based their definition of resistance on the capacity of
drug target genes that could explain variations in drug suscep-
strains to grow in the presence of 0.6 M drug (i.e., 0.15
mg/liter) in a 24-h uracil uptake assay; in that study, sensitive
Several mutations related to resistance to pyrimethamine
strains exhibited Ն50% inhibition at this concentration,
had already been identified in the DHFR-TS gene using site-
whereas resistant strains were less inhibited (Յ35% for mod-
directed mutagenesis and transgenic experiments. Most are
erate resistance and Յ25% for high resistance). Although this
mutations that were known to confer drug resistance in Plas-
narrow range between sensitive and resistant strains can be
modium falciparum. Among these mutations, the T83N muta-
assumed for mutants derived from the same strain (RH) and
tion (S108N in P. falciparum) was found to probably confer
under very stringent experimental conditions, this threshold
resistance to pyrimethamine (15). This resistance is even in-
might not be relevant for clinical isolates, which likely present
creased when this mutation is associated with a mutation of
a higher level of natural diversity. Other authors arbitrarily
S36R and F245S (42). Additional mutations not previously
used an eightfold increase in the inhibitory concentration com-
found in P. falciparum were also produced in T. gondii (W25R,
pared to that of sensitive strains (15). For SDZ and atova-
L98S, and L134H) and were found to generate low levels of
quone, no criterion has been defined, but resistant mutants
resistance (41). Other mutations not related to drug resistance
obtained by mutagenesis have IC s that are 20- to 300-fold
(41, 42) or not affecting the amino acid sequence encoded by
higher than that of the wild type (6, 38, 39).
the DHFR gene have also been described (6); here, two of
Rather than selecting an arbitrary value, we propose a new
these silent mutations were found in two strains (one atypical
approach for a better definition of resistance based on the
distribution of observed IC s. For pyrimethamine and atova-
In the four strains that presented the highest IC s for py-
values of the 16 studied strains follow a log-normal
rimethamine, only silent mutations were found in three strains,
distribution (P ϭ 0.24 and P ϭ 0.11, respectively, by normality
i.e., 156-L in the two type I (B1 and ENT) strains and 145-V in
test). We can estimate that less than 0.1% of strains will have
Resistance of T. gondii to sulfonamide can be induced ex-
tions, i.e., 0.52 mg/liter for pyrimethamine and 0.14 mg/liter for
perimentally and has been related to a mutation at position 407
atovaquone. These concentrations (or higher) can be consid-
in exon 3 of DHPS (40). This mutation was also evidenced on
ered to be unusually high and indicative of resistance. For
a clinical isolate (among 32 human strains) obtained from a
SDZ, IC s did not fit a log-normal distribution (P ϭ 0.03), as
congenitally infected child; it was suggested that this mutation
three strains showed outstandingly high values (IC
was “natural” since no treatment had been administered dur-
mg/liter), possibly synonymous with resistance. A 1% threshold
ing pregnancy and before the isolation of T. gondii (6). This
for abnormally distributed strains can be calculated from the
mutation was not found among the 17 strains studied here.
mean value of “sensitive” strains (excluding the three strains
This was also the case in a study performed on 29 strains
of Ն50 mg/liter) plus 3.3 standard deviations,
isolated from patients with congenital toxoplasmosis (37). In
giving a value of 25.6 mg/liter. Under this condition, none of
our study, various mutations not affecting the amino acid se-
the strains tested in this study were resistant to pyrimethamine
quence encoded by the DHPS gene were found, mainly in the
or atovaquone, and the three strains with IC s for SDZ of Ͼ50
type I and type I-related strains. One resistant strain presented
mg/liter can actually be considered to be resistant to this drug.
mutations found in all type I strains, and one had no detectable
In conclusion, we found some variability in the susceptibili-
mutation. A mutation in exon 5, at position 587, converting
ties of T. gondii strains to pyrimethamine and atovaquone but
alanine to valine was demonstrated for one strain that was
with no clear evidence of drug resistance to these drugs and no
resistant to SDZ. It is noteworthy that this strain was isolated
relationships with strain genotype or defined mutations in drug
from the placenta in a case of congenital toxoplasmosis and
target genes. A higher variability was found for SDZ, with
that prenatal treatment with pyrimethamine and sulfadoxin
possible resistance for three strains. The presence of a new
had been administered for 12 weeks. In this case, the possible
mutation on the DHPS gene of one of these strains could
selection of a drug-resistant mutant cannot be excluded.
possibly be related to this resistance, but such a link needs to
Whether the mutation that we found in this strain can be
be explored by testing additional strains and confirmed by
selected under drug pressure and related to resistance to SDZ
directed mutagenesis experiments. Our proposal of threshold
needs to be investigated experimentally.
concentrations for the definition of in vitro resistance could be
Concerning atovaquone, there is genetic evidence that this
helpful to facilitate the screening for resistance for a large
drugs targets the cytochrome BC1 complex of T. gondii by
number of strains. In addition, the relationship between in
binding to the Q domain of cytochrome b and that mutations
vitro susceptibility and clinical resistance remains to be exam-
within this region are probable mediators of drug resistance
ined. Pharmacokinetic studies have revealed marked individ-
(31, 32). M129L and I254L mutations have been identified
ual variations in pyrimethamine concentrations in human im-
after selection of N-ethyl-N-nitrosourea-mutagenized T. gondii
munodeficiency virus (HIV)-infected patients (20, 49) and in
and related to atovaquone resistance (25, 32). In our study,
children treated for congenital toxoplasmosis (11, 33, 44, 47).
none of these mutations was detected.
In HIV-infected patients with toxoplasmic encephalitis, a pos-
Whether the strains that presented higher IC s for py-
itive relationship between clinical and radiological responses
rimethamine, atovaquone, and SDZ can be considered to be
and median atovaquone plasma concentrations has been
resistant is debatable. In most studies, resistance is arbitrarily
shown (46). Due to these individual variations in drug phar-
defined as a marked decrease in in vitro susceptibility. In a
macokinetics, the differentiation between treatment failures
genetic study of resistance to pyrimethamine, Reynolds et al.
due to insufficient plasma or tissue drug concentration and true
drug resistance should rely on a better documentation of these
20. Jacobson, J. M., M. Davidian, P. M. Rainey, R. Hafner, R. H. Raasch, and
failures, with determinations of plasma levels and attempts to
B. J. Luft. 1996. Pyrimethamine pharmacokinetics in human immunodefi- ciency virus-positive patients seropositive for Toxoplasma gondii. Antimi-
isolate strains in patients under treatment.
crob. Agents Chemother. 40:1360–1365.
21. Katlama, C., B. Mouthon, D. Gourdon, D. Lapierre, F. Rousseau, et al. 1996.
Atovaquone as long-term suppressive therapy for toxoplasmic encephalitis in
ACKNOWLEDGMENTS
patients with AIDS and multiple drug intolerance. AIDS 10:1107–1112.
22. Kaufman, H. E., J. S. Remington, M. L. Melton, and L. Jacobs. 1959.
This study was supported by grants from the Region Champagne-
Relative resistance of slow-growing strains of Toxoplasma gondii to py-
Ardennes (France), the BRC Toxoplasma, and the Centre d’Etudes et
rimethamine. AMA Arch. Ophthalmol. 62:611–615.
23. Kazanjian, P., W. Armstrong, P. A. Hossler, W. Burman, J. Richardson,
We thank all the BRC members who provided T. gondii strains. We
C. H. Lee, L. Crane, J. Katz, and S. R. Meshnick. 2000. Pneumocystis carinii
mutations are associated with duration of sulfa or sulfone prophylaxis expo-
Sulahian for reviewing the manuscript.
sure in AIDS patients. J. Infect. Dis. 182:551–557.
24. Kazanjian, P., W. Armstrong, P. A. Hossler, C. H. Lee, L. Huang, C. B. Beard, J. Carter, L. Crane, J. Duchin, W. Burman, J. Richardson, and S. R. REFERENCES Meshnick. 2001. Pneumocystis carinii cytochrome b mutations are associated
1. Ajzenberg, D., N. Cogne, L. Paris, M. H. Bessieres, P. Thulliez, D. Filisetti,
with atovaquone exposure in patients with AIDS. J. Infect. Dis. 183:819–822. H. Pelloux, P. Marty, and M. L. Darde ´. 2002. Genotype of 86 Toxoplasma
25. Kessl, J. J., K. H. Ha, A. K. Merritt, S. R. Meshnich, and B. L. Trumpower. gondii isolates associated with human congenital toxoplasmosis, and corre-
2006. Molecular basis of Toxoplasma gondii atovaquone resistance modeled
lation with clinical findings. J. Infect. Dis. 186:684–689.
in Saccharomyces cerevisiae. Mol. Biochem. Parasitol. 146:255–258.
2. Ajzenberg, D., A. Dumetre, and M. L. Darde ´. 2005. Multiplex PCR for typing
26. Khan, A., C. Su, M. German, G. A. Storch, D. B. Clifford, and L. D. Sibley. 2005.
strains of Toxoplasma gondii. J. Clin. Microbiol. 43:1940–1943.
Genotyping of Toxoplasma gondii strains from immunocompromised patients
3. Allegra, C. J., J. A. Kovacs, J. C. Drake, J. C. Swan, B. A. Chabner, and H.
reveals high prevalence of type I strains. J. Clin. Microbiol. 43:5881–5887. Masur. 1987. Potent in vitro and in vivo anti Toxoplasma activity of the
27. Khan, A., C. Jordan, C. Muccioli, A. L. Vallochi, L. V. Rizzo, R. Belfort, Jr.,
lipid-soluble antifolate trimetrexate. J. Clin. Investig. 79:478–482. R. W. Vitor, C. Silveira, and L. D. Sibley. 2006. Genetic divergence of
4. Araujo, F. G., J. Huskinson, and J. S. Remington. 1991. Remarkable in vitro Toxoplasma gondii strains associated with ocular toxoplasmosis, Brazil.
and in vivo activities of the hydroxynaphthoquinone 566C80 against
Emerg. Infect. Dis. 12:942–949.
tachyzoites and tissue cysts of Toxoplasma gondii. Antimicrob. Agents Che-
28. Kovacs, J. A., C. J. Allegra, J. Beaver, D. Boarman, M. Lewis, J. E. Parrillo,
mother. 35:293–299. B. Chabner, and H. Masur. 1989. Characterization of de novo folate syn-
5. Araujo, F. G., J. Huskinson-Mark, W. E. Gutteridge, and J. S. Remington.
thesis in Pneumocystis carinii and Toxoplasma gondii: potential for screening
1992. In vitro and in vivo activities of the hydroxynaphthoquinone 566C80
therapeutic agents. J. Infect. Dis. 160:312–320.
against the cyst form of Toxoplasma gondii. Antimicrob. Agents Chemother.
29. Luft, B. J., and J. S. Remington. 1992. Toxoplasmic encephalitis in AIDS. 36:326–330.
Clin. Infect. Dis. 15:211–222.
6. Aspinall, T. V., D. H. Joynson, E. Guy, J. E. Hyde, and P. F. Sims. 2002. The
30. Massimine, K. M., L. T. Doan, C. A. Atreya, T. T. Stedman, K. S. Anderson,
molecular basis of sulfonamide resistance in Toxoplasma gondii and impli-
K. A. Joiner, and I. Coppens. 2005. Toxoplasma gondii is capable of exoge-
cations for the clinical management of toxoplasmosis. J. Infect. Dis. 185:
nous folate transport. A likely expansion of the BT1 family of transmem-
brane proteins. Mol. Biochem. Parasitol. 144:44–54.
7. Baatz, H., A. Mirshahi, J. Puchta, H. Gumbel, and L. O. Hattenbach. 2006.
31. McFadden, D. C., F. Seeber, and J. C. Boothroyd. 1997. Use of Toxoplasma
Reactivation of Toxoplasma retinochoroiditis under atovaquone therapy in
gondii expressing -galactosidase for colorimetric assessment of drug activity
an immunocompetent patient. Ocul. Immunol. Inflamm. 14:185–187.
in vitro. Antimicrob. Agents Chemother. 41:1849–1853.
8. Cassaing, S., M. H. Bessieres, A. Berry, A. Berrebi, R. Fabre, and J. F.
32. McFadden, D. C., S. Tomavo, E. A. Berry, and J. C. Boothroyd. 2000. Magnaval. 2006. Comparison between two amplification sets for molecular
Characterization of cytochrome b from Toxoplasma gondii and Q(o) domain
diagnosis of toxoplasmosis by real-time PCR. J. Clin. Microbiol. 44:720–724.
mutations as a mechanism of atovaquone-resistance. Mol. Biochem. Parasi-
9. Chirgwin, K., R. Hafner, C. Leport, J. Remington, J. Andersen, E. M. Bosler,
tol. 108:1–12. C. Roque, N. Rajicic, V. McAuliffe, P. Morlat, D. T. Jayaweera, J. L. Vilde
33. McLeod, R., D. Mack, R. Foss, K. Boyer, S. Withers, S. Levin, J. Hubbell, B. J. Luft, et al. 2002. Randomized phase II trial of atovaquone with py- and the Toxoplasmosis Study Group. 1992. Levels of pyrimethamine in sera
rimethamine or sulfadiazine for treatment of toxoplasmic encephalitis in
and cerebrospinal and ventricular fluids from infants treated for congenital
patients with acquired immunodeficiency syndrome. Clin. Infect. Dis. 34:
toxoplasmosis. Antimicrob. Agents Chemother. 36:1040–1048.
34. Montoya, J. G., and O. Liesenfeld. 2004. Toxoplasmosis. Lancet 363:1965–1976.
10. Cook, M. K. 1958. The development of a pyrimethamine-resistant line of
35. Nagesha, H. S., S. Din, G. J. Casey, A. I. Susanti, D. J. Fryauff, J. C. Reeder, Toxoplasma under in vitro conditions. Am. J. Trop. Med. 7:400–402. and A. F. Cowman. 2001. Mutations in the pfmdr1, dhfr and dhps genes of
11. Corvaisier, S., B. Charpiat, C. Mounier, M. Wallon, G. Leboucher, M. Al Plasmodium falciparum are associated with in-vivo drug resistance in West
Kurdi, J. F. Chaulet, and F. Peyron. 2004. Population pharmacokinetics of
Papua, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 95:43–49.
pyrimethamine and sulfadoxine in children treated for congenital toxoplas-
36. Petersen, E. 2007. Prevention and treatment of congenital toxoplasmosis.
mosis. Antimicrob. Agents Chemother. 48:3794–3800.
Expert Rev. Anti Infect. Ther. 5:285–293.
12. Dannemann, B., J. A. McCutchan, D. Israelski, D. Antoniskis, C. Leport, B. Luft, J. Nussbaum, N. Clumeck, P. Morlat, J. Chiu, et al. 1992. Treatment
37. Peyron, F., N. Eudes, F. de Monbrison, M. Wallon, and S. Picot. 2004.
of toxoplasmic encephalitis in patients with AIDS. A randomized trial com-
Fitness of Toxoplasma gondii is not related to DHFR single-nucleotide poly-
paring pyrimethamine plus clindamycin to pyrimethamine plus sulfadiazine.
morphism during congenital toxoplasmosis. Int. J. Parasitol. 34:1169–1175.
Ann. Intern. Med. 116:33–43.
38. Pfefferkorn, E. R., S. E. Borotz, and R. F. Nothnagel. 1992. Toxoplasma
13. Derouin, F., and C. Chastang. 1988. Enzyme immunoassay to assess effect of gondii: characterization of a mutant resistant to sulfonamides. Exp. Parasitol.
antimicrobial agents on Toxoplasma gondii in tissue culture. Antimicrob. 74:261–270.
Agents Chemother. 32:303–307.
39. Pfefferkorn, E. R., S. E. Borotz, and R. F. Nothnagel. 1993. Mutants of
14. Derouin, F., and C. Chastang. 1989. In vitro effects of folate inhibitors on Toxoplasma gondii resistant to atovaquone (566C80) or decoquinate. J. Para-
Toxoplasma gondii. Antimicrob. Agents Chemother. 33:1753–1759.
sitol. 79:559–564.
15. Donald, R. G., and D. S. Roos. 1993. Stable molecular transformation of
40. Radke, J. R., B. Striepen, M. N. Guerini, M. E. Jerome, D. S. Roos, and Toxoplasma gondii: a selectable dihydrofolate reductase-thymidylate syn-
M. W. White. 2001. Defining the cell cycle for the tachyzoite stage of Toxo-
thase marker based on drug-resistance mutations in malaria. Proc. Natl. plasma gondii. Mol. Biochem. Parasitol. 115:165–175.
Acad. Sci. USA 90:11703–11707.
41. Reynolds, M. G., J. Oh, and D. S. Roos. 2001. In vitro generation of novel
16. El Hajj, H., M. Lebrun, S. T. Arold, H. Vial, G. Labesse, and J. F.
pyrimethamine resistance mutations in the Toxoplasma gondii dihydrofolate
Dubremetz. 2007. ROP18 is a rhoptry kinase controlling the intracellular
reductase. Antimicrob. Agents Chemother. 45:1271–1277.
proliferation of Toxoplasma gondii. PLoS Pathog. 3:e14.
42. Reynolds, M. G., and D. S. Roos. 1998. A biochemical and genetic model for
17. Grigg, M. E., J. Ganatra, J. C. Boothroyd, and T. P. Margolis. 2001. Unusual
parasite resistance to antifolates. Toxoplasma gondii provides insights into
abundance of atypical strains associated with human ocular toxoplasmosis.
pyrimethamine and cycloguanil resistance in Plasmodium falciparum. J. Biol.
J. Infect. Dis. 184:633–639.
Chem. 273:3461–3469.
18. Harris, C., M. P. Salgo, H. B. Tanowitz, and M. Wittner. 1988. In vitro
43. Romand, S., M. Pudney, and F. Derouin. 1993. In vitro and in vivo activities
assessment of antimicrobial agents against Toxoplasma gondii. J. Infect. Dis.
of the hydroxynaphthoquinone atovaquone alone or combined with py-
157:14–22.
rimethamine, sulfadiazine, clarithromycin, or minocycline against Toxo-
19. Howe, D. K., and L. D. Sibley. 1995. Toxoplasma gondii comprises three plasma gondii. Antimicrob. Agents Chemother. 37:2371–2378.
clonal lineages: correlation of parasite genotype with human disease. J. In-
44. Schmidt, D. R., B. Hogh, O. Andersen, J. Fuchs, H. Fledelius, and E.
fect. Dis. 172:1561–1566. Petersen. 2006. The national neonatal screening programme for congenital TOXOPLASMA DRUG SUSCEPTIBILITY AND RESISTANCE
toxoplasmosis in Denmark: results from the initial four years, 1999-2002. Jaussaud, G. Remy, D. Dupouy, H. Millart, J. M. Pinon, and S. Urien. 2004.
Arch. Dis. Child. 91:661–665.
Population pharmacokinetics of pyrimethamine and sulfadoxine in children
45. Tomavo, S., and J. C. Boothroyd. 1995. Interconnection between organellar
with congenital toxoplasmosis. Br. J. Clin. Pharmacol. 57:735–741.
functions, development and drug resistance in the protozoan parasite Toxo-
48. Tsuchiya, S., M. Yamabe, Y. Yamaguchi, Y. Kobayashi, T. Konno, and K. plasma gondii. Int. J. Parasitol. 25:1293–1299. Tada. 1980. Establishment and characterization of a human acute mono-
46. Torres, R. A., W. Weinberg, J. Stansell, G. Leoung, J. Kovacs, M. Rogers,
cytoid leukemia cell line (THP-1). Int. J. Cancer 26:171–176. J. Scott, et al. 1997. Atovaquone for salvage treatment and suppression of
49. Winstanley, P., S. Khoo, S. Szwandt, G. Edwards, E. Wilkins, J. Tija, R.
toxoplasmic encephalitis in patients with AIDS. Clin. Infect. Dis. 24:422– Coker, W. McKane, N. Beeching, S. Watkin, and A. Breckenridge. 1995.
Marked variation in pyrimethamine disposition in AIDS patients treated for
47. Trenque, T., N. Simon, I. Villena, C. Chemla, C. Quereux, B. Leroux, R.
cerebral toxoplasmosis. J. Antimicrob. Chemother. 36:435–439.
J. Astron. Space Sci. 23(3), 167–176 (2006) THE PEAK ENERGY–DURATION CORRELATION AND POSSIBLE IMPLICATIONS ON GAMMA RAY BURST PROGENITOR Heon-Young Chang Department of Astronomy and Atmospheric Sciences, Kyungpook National University1370 Sankyuk-dong, Buk-gu, Daegu 702-701, Korea (Received June 26, 2006; Accepted July 19, 2006) ABSTRACT We investigate the correlation between the
 inhibitory effect of SDZ increased more progressively for in-creasing concentrations in the cultures (Fig. 2b).
inhibitory effect of SDZ increased more progressively for in-creasing concentrations in the cultures (Fig. 2b).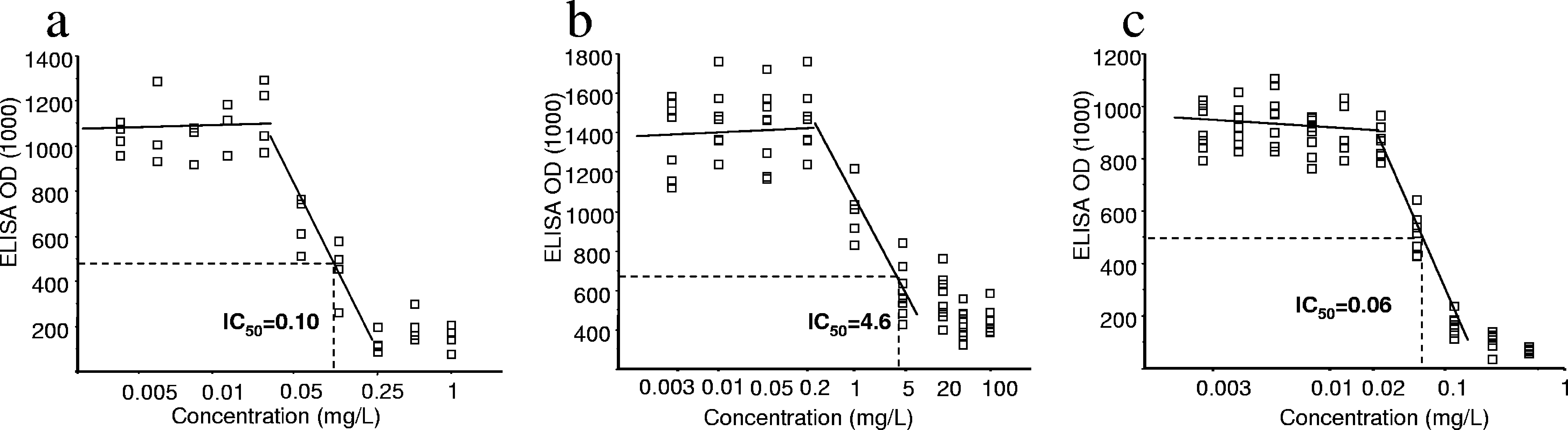
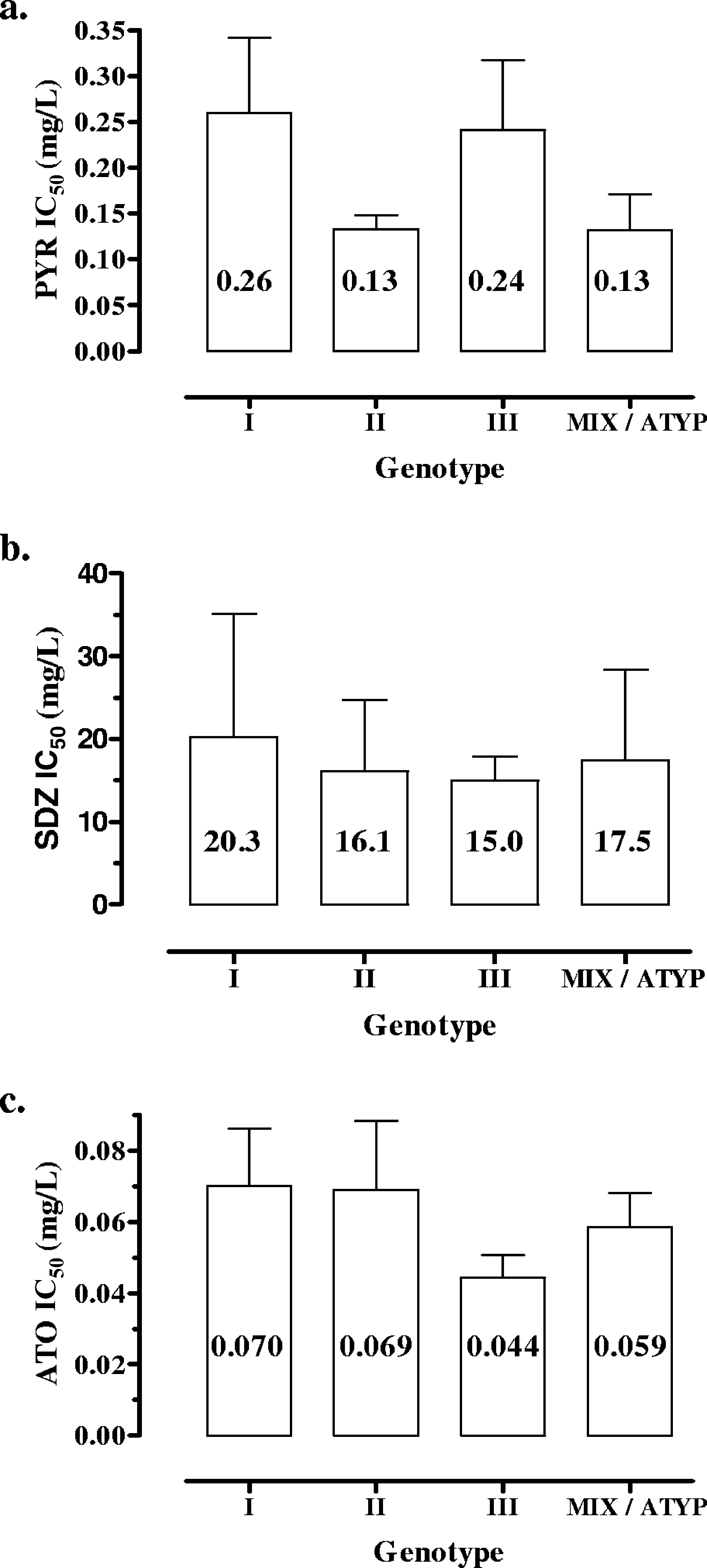
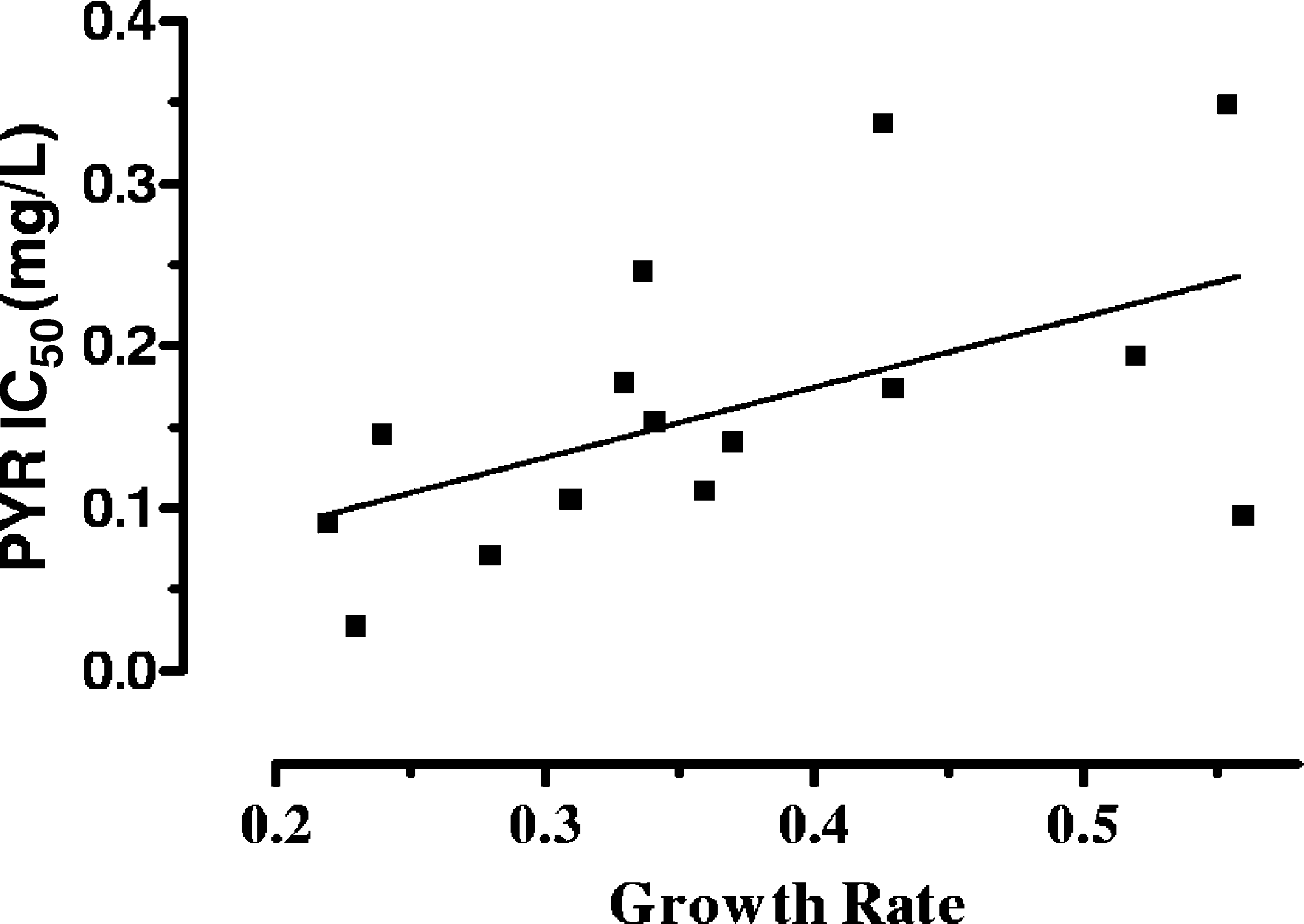 FIG. 2. Representative experiment of in vitro activities of pyrimethamine, SDZ, and atovaquone (strain ME49, type II). Optical density (OD)
values for the ELISA with infected monolayers were plotted versus the concentrations of antimicrobial agents (mg/liter). IC s are estimated from
regression analyses. (a) Pyrimethamine. (b) SDZ. (c) Atovaquone.
FIG. 2. Representative experiment of in vitro activities of pyrimethamine, SDZ, and atovaquone (strain ME49, type II). Optical density (OD)
values for the ELISA with infected monolayers were plotted versus the concentrations of antimicrobial agents (mg/liter). IC s are estimated from
regression analyses. (a) Pyrimethamine. (b) SDZ. (c) Atovaquone.