______________________________________________________________________________ Haloperidol regulates the binding of guanine nucleotides to synaptic membranes through the NMDA receptor N. Natsvlishvili, E. Juravliova, D. Dzneladze and D. Mikeladze* Department of Neurochemistry, I. Beritashvili Institute of Physiology, Georgian Academy of Sciences,14 Gotua St., 380060 Tbilisi, Georgia
The NMDA receptor is believed to be important in a wide range of nervous system functionsincluding neuronal migration, synapse formation, learning and memory. In addition, it isinvolved in excitotoxic neuronal cell death that occurs in a variety of acute and chronicneurological disorders. Besides agonist/coagonist sites, other modulator sites, includingthe butyrophenone site, may regulate the NMDA receptor. It has been shown that manyneuronal modulator mechanisms may be co-coordinated by a group of binding proteinsthat both clusters NMDA receptors and links them to signalling pathways within the cell. We have found that 5'-guanylylimidodiphosphate (Gpp(NH)p) inhibited the binding of[3H]haloperidol to both the cortical and hippocampal synaptic membranes with high affinityand, reciprocally, haloperidol reduced the binding of [3H]Gpp(NH)p to the membranes. Both effects are abolished by addition of anti-p21Ras. Affinity-purified preparations of theNMDA receptor, which were immunoprecipitated by anti-p21Ras contained only the ε2(NR2A/NR2B) subunits of NMDA receptors and nNOS. These data suggest that the possibleproteins participating in the formation of the macromolecular signalling complexes inpostsynaptic density may be nNOS and p21Ras. Keywords: haloperidol, NMDA receptor, nNOS, p21Ras 1. INTRODUCTION
found that a p21Ras-activating protein, SynGAP (GTPaseactivating protein), is exclusively localized at synapses and
The NMDA receptor is believed to be important in a
associated with the PSD95 and NMDA receptors [7, 13].
wide range of nervous system functions including neuronal
It has also been shown that NMDA receptor stimulation
migration, synapse formation and learning and memory.
leads to activation of p21Ras through generation of nitric
In addition, it is involved in the excitotoxic neuronal cell
oxide (NO) in a cGMP-dependent pathway via nNOS [24].
death that occurs in a variety of acute and chronic
These data suggest that p21Ras may also participate in the
neurological disorders (see [21] for a review).
formation of supramolecular complexes with NMDA
It has been shown that many neuronal modulator
mechanisms may be co-coordinated by a group of binding
Besides its agonist/coagonist sites, the NMDA receptor
proteins that both clusters NMDA receptors and links them
is regulated by other modulator action, including voltage-
to signalling pathways within the cell. The NR2A and
dependent blocking of the channel by Mg2+, voltage-
NR2B subunits of the receptor bind to the SH2 domains
independent actions of Zn2+, the redox state of the receptor,
of phospholipase C in a tyrosine phosphorylation-
and sites for arachidonic acid, ethanol, neurosteroids, pH
dependent manner [11]. Members of the PSD-95/SAP90
and polyamines [23]. It has been found that haloperidol, a
family will also cluster NMDA receptors when co-
therapeutically useful antipsychotic drug, inhibits neuronal
expressed in transfected cells and are associated with the
NMDA responses and has neuroprotective effects against
NMDA receptor in neuronal membranes [14,18]. PSD-95
NMDA-induced neurotoxicity [12,19]. Results from
also binds to non-inducible nitric oxide synthase (nNOS)
Whittemore et al. [22] suggest that a noncompetitive
by interaction of its PDZ domain with a PDZ domain in
allosteric modulator site expressed by isoforms of the
nNOS [2] thus suggesting that PDZ domains may
receptor containing the NR1/NR2B subunit mediates
participate in the assembly of macromolecular signalling
haloperidol’s action on the NMDA receptor. The ligand
complexes involving NMDA receptors and one or more
binding experiments of [8], as well as point mutation
signalling molecules at the postsynaptic density.
studies [3] showed that haloperidol interacts with
One of the possible signalling proteins participating in
polyamine sensitive sites of the NR2B subunits.
the formation of the macromolecular signalling complexes
Haloperidol induces the expression of immediate early
in postsynaptic density may be p21Ras [20]. It has been
genes [15] and genes associated with synaptic plasticity [9]. NMDA receptor antagonists diminished the c-fos mRNAincrease produced by haloperidol, indicating that theNMDA receptor is involved in this process [17, 25].
However, the signalling pathway whereby haloperidol
Tel: +99 532 37 47 24; Fax: +99 532 94 10 45;
induces the expression of genes through the
NMDA receptor is unknown. In this study we have tried
Journal of Biological Physics and Chemistry 1 (2001) 24–28 ______________________________________________________________________________ Effects of haloperidol on NMDA-receptor N. Natzvlishvili, E. Juravliova, D. Dzneladze and D. Mikeladze 25
to demonstrate that the action of haloperidol may be
synaptic membranes. It was found that Gpp(NH)p inhibits
mediated by p21Ras, which forms a macromolecular
the binding of 3H-haloperidol to the both cortical
(IC = 1012.8 nM, data not shown) and hippocampal
(IC = 900.6 nM) synaptic membranes with high affinity
(fig.1). On the contrary haloperidol reduces the binding
2. MATERIALS AND METHODS
Membrane preparations from rat cortex or hippo-
campus were obtained after tissue homogenization in 20volumes of ice-cold 0.32 M sucrose, containing 0.1 M
phenylmethylsulphonyl fluoride (PMSF), 3 mM EDTA,5 units/ml aprotinin and 5 µg/ml pepstatin A. The
homogenate was centrifuged at 1000 g for 10 min, the pelletwas removed, and the supernatant was centrifuged at 20000
g for 20 min. The pellet was resuspended in 20 mM Tris-
HCl, pH 7.4 buffer, containing 0.1 mM PMSF,
2 mM EDTA, 5 µg/ml pepstatin and 5 units/ml aprotinin
(buffer A) to yield a suspension of 5 mg protein/ml and
Solubilization was carried out using 1% sodium deoxy-
cholate at a detergent/protein ratio = 4/1 (mg/mg) in buffer
A during 1 hour at 4 °C followed by centrifugation at
100000 g for 1 hour. The final supernatant was dialyzedagainst 20 mM Tris-HCl (pH 7.4) containing 2 mM EDTA
Figure 1. Inhibition of 5 nM [3H]haloperidol binding by GppNHp
and 0.1 mM PMSF (buffer B) and was applied on either
in rat hippocampal synaptic membranes in the presence (•) and
dextrorphan-Sepharose or trihexyphenidyl (THP)-Sepha-
absence (!) of anti-Ras. 100 % binding corresponds to the specific
rose columns (1 x 10 cm) pre-equilibrated in buffer A.
binding of the control in the absence of GppNHp. Binding wascarried out as described in the experimental procedures. The curve
The columns were washed with 40 ml buffer A and
is representative of three independent experiments. Specific
matrix-binding proteins were eluted by 100 mM dex-
binding in the absence of GppNHp (dpm ± S.E.M.) from
trorphan or by 100 mM phencyclidine in buffer A. The
eluates were dialyzed against buffer B and their bindingactivity was determined.
[3H]haloperidol binding to synaptic membranes, and
the solubilized or affinity-purified preparations, weredetermined in buffer A, containing 200–300 µg/ml protein
and 5 nM [3H]haloperidol. Nonspecific binding wascalculated after addition 0.5 mM of haloperidol to the
medium. The incubation was carried out during 1 hour at
4 oC and the mixture was filtered through Whatman GF/B
filters pretreated with 0.05 polyethylenimine. [3H]Gpp(NH)p binding assays were carried out in the
presence of 5 nM [3H]Gpp(NH)p as described for[3H]haloperidol binding. 1 mM Gpp(NH)p was used to
define the nonspecific binding. Radioactivity retained on
the filter was determined by liquid scintillation
The binding of THP and dextrorphan to Sepharose 4B
was carried out as Egly et al. [10] suggested for
triflufenazine immobilization [6]. Protein concentration
was determined using a dye-binding method (Bio-Rad).
Figure 2. Inhibition of 5 nM [3H]GppNHp binding byhaloperidol in rat hippocampal synaptic membranes in thepresence (•) and absence (!) of anti-Ras. 100 % binding
3.RESULTS
corresponds to the specific binding of the control in the absenceof haloperidol. Binding was carried out as described in the
3.1 Binding experiments
experimental procedures. The curve is representative of threeindependent experiments. Specific binding in the absence of
The guanine nucleotide analogue, 5'-guanylyli-
haloperidol (dpm ± S.E.M.) from [3H]GppNHp was 2773 ± 248.
midodiphosphate (Gpp(NH)p), was examined for its effecton 3H-haloperidol binding to rat cortical and hippocampal
______________________________________________________________________________
26 N. Natzvlishvili, E. Juravliova, D. Dzneladze and D. Mikeladze Effects of haloperidol on NMDA-receptor
of [3H] Gpp(NH)p to the membranes (fig. 2). Both effects
solubilization by 1% sodium deoxycholate (see Materials
are abolished by addition of anti-p21Ras.
and Methods) and subsequent centrifugation was applied
In the next series of experiments the effects of other
either on THP-Sepharose or dextrorphan-sepharose
NMDA receptor and sigma antagonists on the binding of
columns. Proteins binding THP-Sepharose or dextrorphan-
[3H] Gpp(NH)p to the hippocampal synaptic membranes
Sepharose from columns were eluted by 10 µM phen-
were examined. We found that out of the series of NMDA
cyclidine or by 10 µM dextrorphan respectively.
antagonists tested, haloperidol is significantly more potent
SDS-PAG electrophoresis has shown that eluates from
at decreasing [3H]Gpp(NH)p binding. Phencyclidine
THP-sepharose (preparation 1) contain 5 major protein
(PCP),MK-801((+)-5-methyl-10,11-dihydro-5H-
fractions with molecular weights of 23000, 48000, 56000,
dibenzo[a,d]cyclohepten-5,10-imine) and dextrorphan
11500 and 125000, and eluates from dextrorphan-
were less potent than haloperidol (Table 1). These data
sepharose (preparation 2) contain 6 protein fractions with
demonstrate that the action of haloperidol on the binding
molecular weights of 23000, 32000, 48000, 56000, 11500,125000 (data not shown). As the dextrorphan has higheraffinity to a sigma receptor, is possible that the additional
Table 1. The inhibition of [3H]GppNHp bindinga to synaptic
protein in the preparation 2 is one of the sigma binding
membranes, and solubilized and affinity-purified preparations.
Pharmacological specificities of solubilized and both
affinity-purified preparations were studied by competitionbinding experiments. MK-801 was found to have the
highest affinity for the proteins of solubilized preparations,
while pentazocine, a sigma opiate agonist, the lowest (data
not shown). This shows that sodium deoxycholate
solubilizes NMDA-glutamate receptors highly effectively,but sigma-binding proteins relatively ineffectively. The
pharmacological profile of the affinity-purified prepa-rations does not significantly differ by specificity from the
solubilized preparations, indicating that the pharmaco-
logical specificity of NMDA receptors does not changeduring affinity chromatography.
It was also revealed that both preparation 1 and
preparation 2 bound [3H] GppNp and did not lose
sensitivity to haloperidol. The data for haloperidol are
presented in Table 1; similar results are obtained for theother drugs. In spite of the fact that the IC is in this case
a100 % binding corresponds to the specific binding of 5nM
lower against the membranous preparation, the specificity
[3H]GppNHp in the absence of drugs. Binding was measured as
of interaction with the drugs is the same (data not shown).
described in the experimental procedures. Specific binding in
Thus it is possible to conclude that the supramolecular
the absence of drugs (dpm ± S.E.M.) from [3H]GppNHp was
complex of NMDA- receptor after affinity chromatography
2773 ± 248 for membranes, 946±85 for the solubilized
contains proteins binding in the same way as haloperidol
preparation and 234± 37 for the affinity-purified preparation. b
Mean ± SEM calculated from 3 independent experiments. 3.3 Immunoprecipitation of the NMDA receptor
of guanine nucleotide with synaptic membranes is specificand can be directed at a system of p21Ras.
For identification of proteins in the supramolecular
complex of NMDA receptor both affinity-purified
3.2 Purification and characterization of haloperidol
preparations were immunoprecipitated by anti-Ras and
binding proteins
analyzed by Western-blot. It was found that only the
ε2 (NR2A/NR2B) subunits of the NMDA receptors were
It has been shown previously that many neuronal
present in both immunoprecipitated preparations. In
modulator mechanisms may be co-coordinated by a group
addition it was revealed that both preparations contained
of binding proteins that both clusters NMDA receptors
nNOS and did not contain the ε1 subunit of the NMDA
and links them to signalling pathways within the cell [21].
The possible signalling proteins participating in the
Thus, it is possible to conclude that the supramolecular
formation of the macromolecular signalling complexes in
complex obtained after dextrorphan-Sepharose and THP-
postsynaptic density may be nNOS and p21Ras [20]. For
Sepharose chromatography contains the NR2A/NR2B
identification of the target protein for haloperidol in the
subunits of the NMDA receptor, p21Ras and nNOS.
supramolecular complex of NMDA receptors the
Furthermore this macromolecular complex is sensitive to
solubilization and affinity chromatography was performed.
haloperidol and binds guanine nucleotides.
For this purpose, the final supernatant followed
______________________________________________________________________________ Effects of haloperidol on NMDA-receptor N. Natzvlishvili, E. Juravliova, D. Dzneladze and D. Mikeladze 27
mediated by the haloperidol/ifenprodil binding sites of theNMDA receptor. Accordingly, it is possible to concludethat regulation of the Ras-signalling pathway via theNMDA receptor may be carried out by polyamine(ifenprodil) sites.
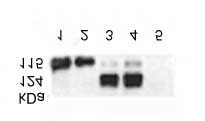
Figure 3. Co-immunoprecipitation of NR2A/NR2B subunits
REFERENCES
of NMDA-receptor and nNOS by anti-Ras. Preparations 1 and 2were precipitated using the Ras antibody, separated on
1. Brenman, J.E., Chao, D.S., Gee, S.H., et al. Interaction
SDS-PAGE, and analyzed using immunoblots probed with
of nitric oxid synthase with the post synaptic density
nNOS, NR2A/NR2B subunits and SAP antibodies as desribed
protein PSD-95 and alpa-1-syntrophin mediated by
in “Materials and Methods”. Lanes 1 and 2 – nNOS in prepara-
PDZ domains. Cell 84 (1996) 757–767.
tions 1 and 2 respectively; lanes 3 and 4 – NR2A/NR2B in prepa-
2. Brenman, J.E., Christopherson, K.S., Graven, S., et al.
rations 1 and 2 respectively; lane 5 – SAP(PSD-95). The blot isrepresentative of three independent experiments.
Cloning and characterization of post synaptic density 93, a nitric oxide synthase interacting protein. J.Neurosci.16 (1996) 7407–7415.
3. Brimecombe, J.C., Gallagher, M.J., Lynch, D.R.,
4. DISCUSSION
Aizenman, E. An NR2B point mutation affectinghaloperidol and CP101, 606 sensitivity of single
Several PDZ-containing proteins have been shown to
recombinant N-methyl-D-aspartate receptors.
interact closely with ion-channel linked receptors (see for
J. Pharmacol. Exp. Therapeutics286 (1998) 627–34.
review [5]). PSD-95 and PSD-93 were reported to interact
4. Boegman, R.J., Vincent, S.R. Involvement of adenosine
directly with the C-terminal domains of two different
and glutamate receptors in the induction of c-fos in the
subunits (NR2A and NR2B) of the NMDA receptor [1,14].
striatum by haloperidol. Synapse22 (1996) 70–77.
The multivalent PDZ-containing protein CIPP (channel
5. Carner, C.C., Nash, J., Huganir, R.L. PDZ domains in
interacting PDZ-domain protein) was shown to bind to all
synapse assembly and signaling. Trends Cell. Biol.10
four NR2 subunits [16]. On the other hand nNOS and
synaptic Ras-GTPase activating protein (synGap),which
6. Charbonneau, H., Porath, J. Purification of plant
have been reported [7,13] to interact with PDZ-containing
calmodulin by flufenazin-sepharose affinity chromato-
proteins, could further contribute to a complex protein
graphy. Biochem. Biophys. Res. Comm.90 (1979)
scaffold. Our data has shown that the affinity-purified
NMDA receptor preparation was immunoprecipitated by
7. Chen, H.J., Rojas-Soto, M., Oquni, A., Kennedy, M.B.
anti-Ras and contained nNOS. The protein complex does
A synaptic Ras-GTPase activating protein (p135
not contain SAP, however, and this fact indicates that the
SynGAP) inhibited by CaM kinase II. Neuron 20
interaction of Ras and nNOS with the NR2B subunit is
possibly carried out by other proteins, such as PSD-93
8. Coughenour, L.L., Cordon, J.J. Characterization of
haloperidol and trifluperidol as subtype-selective N-
Ras signalling at the postsynaptic membrane may be
methyl-D-aspartate receptor antagonists using [3H]TCP
involved in the modulation of excitatory synaptic
and [3H]Ifenprodil binding in rat brain membranes.
transmission [20]. On the basis of our results it is possible
J. Pharmacol. Exp. Therapeutics 280 (1997) 584–592.
to conclude that the macromolecular complex containing
9. Eastwood, S.I., Heffernan, J., Harrison, P.J. Chronic
NR2B and Ras may be directly regulated by haloperidol.
haloperidol treatment affects the expression of synaptic
As shown in fig. 1, haloperidol decreases the binding of
and neuronal plasticity-associated genes. Mol.
3H-GppNp with synaptic membranes in a dose-dependent
Psychiatry2 (1997) 322–329.
manner. Neither PCP, MK-801, dextrorphan nor penta-
10. Egly, J.-M., Porath, J. Change transfer and water
zocine change the binding of 3H-GppNp with synaptic
mediated chromatography. II. Adsorption of nucleo-
membranes, which suggests that haloperidol binds
tides and related compounds in acriflavin-Sephadex.
elsewhere than at the PCP-site of the NMDA receptor. Ilyin
J. Chromatogr.168 (1979) 35–47.
et al. [12] indicated that haloperidol selectively inhibits
11. Gurd, J.W., Bisson, N. The N-methyl-d-aspartate
NMDA receptors composed of NR1 and NR2B subunits
receptor subunits NR2A and NR2B bind to the
by acting at a noncompetitive allosteric site distinct from
SH2-domains of phospholipase C. J. Neurochem. 69
the glutamate-, glycine- and PCP-binding sites, but related
to the ifenprodil binding sites. Brimecombe et al. [3] found
12. Ilyin, V.I., Whittemore, E.R., Guastella, J., et al.
an NR2B point mutation affecting the haloperidol and
Subtype-specific inhibition of NMDA receptors by
CP101 (ifenprodil analogue) sensitivity of a single
haloperidol. Mol. Pharmacol, 50 (1996) 1541–1550.
recombinant NMDA receptor. Together, these observations
13. Kim, J.H., Liao, D., Lau, L.F., Huganir, R.L. SynGAP:
suggest that the unique effect of haloperidol on 3H-GppHp
a synaptic RasGAP that associates with the PSD-95/
binding (and possibly on the activity of p21Ras) may be
SAP90 protein family. Neuron20 (1998) 683–691. ______________________________________________________________________________
28 N. Natzvlishvili, E. Juravliova, D. Dzneladze and D. Mikeladze Effects of haloperidol on NMDA-receptor
14. Kornau, H.-C., Schenker, L.T., Kennedy, M.B.,
22. Whittemore, E.R., Ilyin, V.I., Woodward, R.M.
Seeburg, P.H. Domain interaction between NMDA
Antagonism of N-methyl-D-aspartate receptors by σ-
receptor subunits and the postsynaptic density protein
site ligands: potency, subtype selectivity and
PSD-95. Science269 (1995) 1737–1740.
mechanisms of inhibition. J. Pharmacol. Exp.
15. Kouradi, C., Heckers, S. Haloperidol-induced Fos
Therapeutics282 (1997) 326–338.
expression in striatum is dependent upon trancription
23. Yamakura, T., Shimoji, K. Subunit- and site-specific
factor cyclic AMP response element binding protein.
pharmacology of the NMDA receptor channel. Neuroscience 65 (1995) 1051–1061. Prog. Neurobiology59 (1999) 279–298.
16. Kurschner, C., Mermelstein, P.G., Holden, W.T.,
24. Yun, H-Y., Gonzalez-Zulueta, M., Dawson, V.L.,
Surmeier, D.J. CIPP, a novel multivalent PDZ domain
Dawson, T.M. Nitric oxide mediates N-methyl-D-
protein, selectively interacts with kir4.0 family
aspartate receptor-induced activation of p21ras.
members, NMDA receptor subunits, neurexins, and
Proc. Natl Acad. Sci. 95 (1998) 5773–5778.
neurogulins. Mol. Cell Neurosci. 11 (1998) 161–172.
25. Ziolkowska, B., Hollt, V. The NMDA receptor
17. Morinobu, S., Strausbaugh, H., Terwilliger, R.,
antagonist MK-801 markedly reduces the induction
Duman, R.S. Regulation of c-fos and NGF-A by
of c-fos gene by haloperidol in the mouse striatum.
antidepressant treatment. Synapse 25 (1997) 313– Neurosci. Lett.156 (1993) 39–42.
18. Neithammer, M., Kim, E., Sheng, M. Interaction
between the C terminus of NMDA receptor subunits
APPENDIX
and multiple members of the PSD-95 family ofmembrane-associated guanylate kinases. J. Neurosci.Alphabetical list of abbreviations: 16 (1996) 2157–2163.
19. Nishikawa, H., Hashino, A., Kume, R., et al. Gpp(NH)p: 5’guanylylimidodiphosphate;
Involvement of direct inhibition of NMDA receptors
EDTA: ethylenediaminetetraacetic acid;
in the effects of -receptor ligands on glutamate neuroto-
MK-80: (+)-5-methyl-10,11-dihydro-5H-dibenzo-
xicity in vitro. Eur.J. Pharmacol. 404 (2000) 41–48.
20. Orban, P.C., Chapman, P.F., Brambilla, R. Is the
NMDA: N-methyl-D-aspartate;
Ras-MAPkinase signalling pathway necessary for
nNOS: neural nitric oxide synthase;
long-term memory formation? Trends Neurosci.22 PMSF: phenylmethylsulphonylfluoride; (1999) 38–44. PSD-95: postsynaptic density protein;
21. Ozawa, S., Kamiya, H., Tsuzuki, K. Glutamate
SAP-90: synapse associated protein;
receptors in the mammalian central nervous system. THP: trihexyphenidylhydrochloride. Prog. Neurobiol.54 (1998) 581–618.
110 CMR: DEPARTMENT OF CHILDREN AND FAMILIES 110 CMR 2.00: GLOSSARY Whenever used throughout 110 CMR, the following words shall have the following meanings, unless the context plainly requires otherwise. Abuse means the non-accidental commission of any act by a caretaker upon a child under age 18 which causes, or creates a substantial risk of physical or emotional injury, or constitutes a sexual
The Frank J. Remington Center University of Wisconsin Law School Greetings! It is a pleasure to present the August edition of ournewsletter to friends and graduates of the Remington Center. As befitsa newsletter coming on the heels of our busy summer program, thisedition is packed with essays by students and clinical faculty, describ-ing the many and varied activities in our clinical progra

 ______________________________________________________________________________
______________________________________________________________________________ ______________________________________________________________________________
______________________________________________________________________________