Das pharmakologische Profil von Sildenafil zeigt neben der PDE5-Inhibition auch eine geringe Aktivität an der PDE6 in der Retina. Dies erklärt visuelle Nebenwirkungen wie Farbsehstörungen, die gelegentlich auftreten. Die orale Bioverfügbarkeit beträgt etwa 40 %, mit einer hohen Bindung an Plasmaproteine. Das Verteilungsvolumen ist groß, sodass die Substanz rasch in verschiedene Gewebe gelangt. Die Metabolisierung erfolgt hepatisch und produziert einen aktiven Metaboliten, der die pharmakologische Wirkung ergänzt. Nebenwirkungen sind dosisabhängig und umfassen Kopfschmerzen, Hautrötung und Dyspepsie. Bei Vergleichen innerhalb der Wirkstoffklasse wird viagra original regelmäßig als Beispiel für eine Substanz mit schneller, aber kurzzeitiger Wirkung aufgeführt.
Ianr.org.cn
Cell Medicine, Part B of Cell Transplantation, Vol. 1, pp. 15–46, 2010
Printed in the USA. All rights reserved.
Copyright 2010 Cognizant Comm. Corp. Cell Therapy From Bench to Bedside Translation in CNS Neurorestoratology Era
Hongyun Huang,* Lin Chen,* and Paul Sanberg†
*Center for Neurorestoratology, Beijing Rehabilitation Center, Beijing, P.R. China
†Department of Neurosurgery, University of South Florida, Tampa, FL, USA
Recent advances in cell biology, neural injury and repair, and the progress towards development of neurorest-orative interventions are the basis for increased optimism. Based on the complexity of the processes ofdemyelination and remyelination, degeneration and regeneration, damage and repair, functional loss andrecovery, it would be expected that effective therapeutic approaches will require a combination of strategiesencompassing neuroplasticity, immunomodulation, neuroprotection, neurorepair, neuroreplacement, and neu-romodulation. Cell-based restorative treatment has become a new trend, and increasing data worldwide havestrongly proven that it has a pivotal therapeutic value in CNS disease. Moreover, functional neurorestorationhas been achieved to a certain extent in the CNS clinically. Up to now, the cells successfully used inpreclinical experiments and/or clinical trial/treatment include fetal/embryonic brain and spinal cord tissue,stem cells (embryonic stem cells, neural stem/progenitor cells, hematopoietic stem cells, adipose-derivedadult stem/precursor cells, skin-derived precursor, induced pluripotent stem cells), glial cells (Schwann cells,oligodendrocyte, olfactory ensheathing cells, astrocytes, microglia, tanycytes), neuronal cells (various pheno-typic neurons and Purkinje cells), mesenchymal stromal cells originating from bone marrow, umbilical cord,and umbilical cord blood, epithelial cells derived from the layer of retina and amnion, menstrual blood-derived stem cells, Sertoli cells, and active macrophages, etc. Proof-of-concept indicates that we have nowentered a new era in neurorestoratology.
Key words: Cell therapy; Translational medicine; Neurorestoratology; Central nervous system diseases
BRIEF PROFILE OF NEURORESTORATOLOGY
neurodegeneration, cerebrovascular anoxia or ischemia,edema, demyelination, sensory and motor disorders, and
neuropathic pain, as well as neural damage resulting
Neurorestoratology, a distinct discipline within the
from toxic, physical, and chemical factors, immune, in-
neurosciences, has been clearly defined by the Interna-
fectious, inflammatory, hereditary, congenital, develop-
tional Association of Neurorestoratology as one subdis-
mental, and other intractable neural lesions (376).
cipline and one new branch of neuroscience, which stud-
Inexorable Law of Neuroscientific Innovation
ies the therapeutic strategies for neural regeneration, repair,and replacement of damaged components of the nervous
Thousands of years ago (approx 2500 B.C.), spinal
system, neuroplasticity, neuroprotection, neuromodula-
cord injuries were described as “crushed vertebra in his
tion, angiogenesis, immunomodulation, and their mech-
neck” as well as symptoms of neurological deterioration
anisms to cause improvement. The core of neurorestora-
without treatment in the ancient Egyptian medical papy-
tology is to restore neurological function in humans. The
rus known as the Edwin Smith Surgical Papyrus by the
research field of neurorestoratology covers various neur-
physician and architect of the Sakkara pyramids Imhotep
orestorative treatments including transplantation of tis-
(8). Nearly 90 years ago Ramon y Cajal (1926) stated
sue and cells, biomaterials and bioengineering, neuro-
with certainty: “Once the development was ended, the
modulation by electrical and/or magnetic stimulation,
founts of growth and regeneration of the axons and den-
pharmaceutical or chemical therapies in neurotrauma,
drites dried up irrevocably. In the adult centers, the
Received February 19, 2010; final acceptance March 22, 2010. Address correspondence to Hongyun Huang, Center for neurorestoratology, Beijing Rehabilitation Center, Beijing, 100144, P.R. China. Tel: 86-10-5882-3400; Fax: 86-10-5162-5950; E-mail: [email protected]
nerve paths are something fixed, ended, and immutable.
potential to restore lost function. Intervention strategies
Everything may die, nothing may be regenerated. It is
include gene therapy, neurotrophic factors, and cell
for the science of the future to change, if possible, this
transplantation (195,248,321,377), tissue engineering
harsh decree” (51). Regeneration and restoration of the
(402), and neurostimulation (172,369).
central nervous system (CNS) was thought to be almost
Neurorestorative Therapy or Surgery. Intervention
an impossibility at that time, although scientists still
strategies include cell-based and pharmacological thera-
tried to study the special mystery of human life through
pies (423) and electrostimulation (373).
transplanting brain tissues (374), electrical stimulation
Restorative Neuroscience. Intervention strategies in-
(156), nerve growth factor (NGF) administration (204),
clude cell transplantation, stimulation, and medicine (11).
Together, all of the medical terms mentioned above
Commonly, physicians of traditional clinical disci-
were not considered as distinct disciplines, but instead a
plines have believed that sequelae of stroke, CNS
branch of neurology, neurosurgery, or a specific kind of
trauma, neurodegenerative diseases, and damage lack ef-
therapy. The emergence of the term “neurorestoratol-
fective treatments. The majority of the medical commu-
ogy,” however, signals the birth of a new discipline,
nity now still think that: “Our knowledge of the patho-
which is equally important in comparison with neurol-
physiological processes, both the primary as well as the
ogy and neurosurgery (59). The potential mechanisms of
secondary, has increased tremendously. However, all
action of neurorestoratology techniques are highlighted
this knowledge has only led to improved medical care
but not to any therapeutic methods to restore, even par-tially, the neurological function” (8). Unfortunately, the
NEURORESTORATIVE MECHANISMS
majority of physicians remain ignorant or unaware of
OF CELL THERAPY
the increasing quantity of published evidence concern-
Neuroprotection by Neurotrophins and Immune
ing CNS functional restoration by neuromodulation,
neuroprotection, axon sprouting, neural circuit recon-struction, neurogenesis, neuroregeneration, neurorepair,
Bone marrow mesenchymal stem cells (BM-MSCs)
and neuroreplacement in animal models and patients
have the capacity to modulate immune/inflammatory re-
sponses in Alzheimer disease (AD) mice, ameliorating
It is the eternal desire for humans to prolong and im-
their pathophysiology, and improving the cognitive de-
prove their quality of life, which, with no doubt, it
cline associated with amyloid-β (Aβ) deposits (199).
should be the instinct and responsibility of physicians
Neural stem/precursor cells (NSPCs) can be used as an
and neuroscientists to search for effective methods. Ob-
immune regulatory tool for autoimmune encephalomy-
viously, it is inappropriate and overly pessimistic for
elitis (300). In parallel, olfactory ensheathing cells (OECs)
physicians to always say that there is no way to help
play a role in neuroprotection through the secretion of
patients with the sequelae of diseases and damage to the
neurotrophins or growth factors (350). Studies by Chopp
CNS. Therefore, the new discipline, Neurorestoratology,
and colleagues have proven that transplanting MSCs
is bound to arise from neuroscientific innovation, filling
into the brain leads to secretion of neurotrophins, growth
in the question-marked frame shown in Figure 1, which
factors and other supportive substances after brain injury
aims to create effective therapeutic strategies to benefit
(307), which change the microenvironment in the dam-
aged area and continually facilitate endogenous neuro-restorative mechanisms by reducing apoptotic cell death
(55). Garbuzova-Davis et al. (114) and Zwart et al. (429)reported that an appropriate dose of mononuclear human
Restorative Neurology. Dimitrijevic put forward this
umbilical cord blood (MNC hUCB) cells may provide a
term in 1985. It was defined as the branch of the neuro-
neuroprotective effect for motor and optic neurons
logical sciences that applied active procedures to im-
through the active involvement of these cells in modu-
prove the functioning of the impaired nervous system
lating the host’s immune inflammatory system response.
through selective structural and functional modification
Neural protection afforded by adipose-derived stromal
of abnormal neurocontrol according to underlying mech-
cells was found to be mostly attributable to activated
anisms and clinically unrecognized residual functions;
caspase-3 and Akt-mediated neuroprotective pathway
its intervention strategies included neurostimulation,
signaling through paracrine support provided by trophic
neuromodulation, and neuroectomies (83), cell trans-
plants (242), drugs (222), and so on. Restorative Neurosurgery. This term was put for-
ward by Liberson in 1987 (221). Then restorative neuro-
In early animal studies using neural stem cell treat-
surgery, as the frontier of neurosurgery, provided the
ments, very few cells become neurons (53) and it was
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Figure 1. Distribution of disease treated by clinical disciplines of neuroscience and relevant cross- disciplines.
believed that there was no evidence that “new” neurons
neuronal subtypes, establish synaptic contact with host
could reinnervate muscle (256). More studies now indi-
cells, increase the expression of synaptic markers, and
cate that mesenchymal stromal cells (137,206) and neu-
enhance axonal reorganization in the injured area. Initial
ral stem cells could survive, migrate, and differentiate
patch-clamp recording demonstrated that the MGE cells
into endothelial cells or glia and neurons (173,427),
received postsynaptic currents from the host cells. Func-
form electrically active and functionally connected neu-
tional recovery could be mediated by neurotrophic sup-
rons (15) that form synapses between host and donor
port, new synaptic circuit elaboration, and enhancement
cells (396), and elicit further functional repair following
of the stroke-induced neuroplasticity (74). Recent find-
transplantation into the adult CNS (81,100,161,210,252,
ings of transplanted embryonic dopamine (DA) neurons
349,382). Furthermore, evidence shows that bone mar-
into the substantia nigra (SN) indicate that DA neurons
row stromal cells are capable of remodeling the blood
could extend neurites towards a desired target through
the brain stem and caudal diencephalon to reconstructthe neural circuitry from grafted neurons in the host
(315,355). Application of stem cells for neuroreplace-
Many different cell types have been shown to have
ment therapy is therefore no longer science fiction—it
potential in the repair or remyelination of CNS diseases
is science fact (367). OECs can promote neuroplasticity
in neurodegenerative diseases (62), while NSCs can sup-press abnormal mossy fiber sprouting into the inner mo-
lecular layer with subsequent reduction of hippocampal
Schwann cells (SCs) can induce sprouting of motor
and sensory axons in the adult rat spinal cord (214). Ac-cumulated studies show that OECs are capable of aiding
Neuromodulation or Unmasking and Signaling Repair
axon growth or sprouting following transplantation and
continued regeneration of the denervated caudal host
Our clinical study showed that patients with chronic
tract resulting in the recovery of neurological functions
spinal cord injury have rapid recovery of some functions
in acute (44,211,312,314,409) as well as chronic spinal
following OEC transplantation. The explanation is that
they changed the local microenvironment by the secre-
Neural Circuit or Network Reconstruction,
tion of useful chemicals or growth factors, which can
Neuroplasticity, and Neuroreplacement
promote the nerve cell growth, unmasking the quiescent
In the rat stroke model, a graft of medial ganglionic
axons, and therefore restoring some of the lost functions
eminence (MGE) cells may differentiate into multiple
(147). Grafting dental pulp stem/stromal cells (DPSC)
can promote proliferation, cell recruitment, and matura-
alternative therapy for a variety of degenerative and
tion of endogenous stem/progenitor cells by modulating
traumatic disorders (Table 1). It has been argued that
the local microenvironment through enhancing ciliary
neural transplantation can promote functional recovery
neurotrophic factor (CNTF), vascular endothelial growth
by the replacement of damaged nerve cells, the reestab-
factor (VEGF), and fibroblast growth factor (FGF) as
lishment of specific nerve pathways lost as a result of
stimulators and modulators of the local repair response
injury, the release of specific neurotransmitters, or the
in the CNS (142) and neuromodulation would likely be
production of factors that promote neuronal growth
necessary to realize the full potential of NSC grafts in
restoring function (111). Another study also suggested
Fetal/Embryonic Spinal Cord Tissue
that the inhibitory neurotransmitters γ-aminobutyric acid(GABA) and glycine secreted by transplanted cells
The first successful transplantation of fetal spinal
could be an effective clinical tool for treating spinal cord
cord into adult spinal cord was reported in 1983 (291).
injury (SCI)-associated neuropathic pain (91).
Some topographical features of the normal spinal cord
Generally, the patient’s functional restoration origi-
may be represented in mature spinal cord transplants
nated from some or all of the mechanisms as listed
(318). Embryonic spinal cord transplants are capable of
above. But under many conditions, functional recovery
replacing damaged intraspinal neuronal populations and
is from neuromodulation or unmasking, neuroprotection,
restoring some degree of anatomical continuity between
sprouting, neural circuit reconstruction, and neuroplas-
the isolated rostral and caudal stumps of the injured
ticity by neurotrophins, immune or inflammatory modu-
mammalian spinal cord (317). Improved hind limb be-
lation and local microenvironment change, and in a few
havioral deficits were observed after fetal spinal cord
cases from neurogenesis or neuroregeneration, and neur-
homografts (31). Moreover, the grafted fetal/embryonic
orepair (378,409). Neuroreplacement may be an impor-
tissue may stimulate partial regression of an established
tant tool for Parkinson’s disease (PD), but may not be a
glial scar (141), replace missing motoneurons (353), and
major way for functional neurorestoration in most other
form myelin (324). Data have shown that spinal cord
CNS diseases or damage. It remains unclear how to ex-
transplants support regrowth of adult host axons result-
plain the exact mechanisms for clinical functional recov-
ing in reconstitution of synaptic complexes within the
ery; in the future mechanisms by which cell transplanta-
transplant that in many respects resemble normal syn-
tion enhances functional recovery need to be better
apses. Transplants of fetal spinal cord may also contrib-
understood following further experimental study.
ute to behavioral recovery by rescuing axotomized hostneurons that otherwise would have died. Electrophysio-
PRECLINICAL STUDIES OF CELL-BASED
logical investigations of functional recovery after intras-
NEURORESTORATOLOGY IN CNS DISEASES
pinal transplantation have been recorded (32,375). Olfactory Ensheathing/Precursor Cells
In 1977, the first evidence was presented that grafts
of fetal brain tissue to the adult CNS could counteract an
OECs are cells that display Schwann cell or astro-
experimentally induced neurological deficit (279). Cell/
cyte-like properties. They are a source of growth factors
tissue suspensions, dissociated from selected embryonic
and adhesion molecules that play a very important role
brain regions, can mediate considerable reinnervation of
as a neuronal support enhancing cellular survival (179,
a previously denervated brain or spinal cord region, and
292). Transplants of these cells have been shown to have
they can replace neurons intrinsic to a particular target
a neuroprotective effect, supporting axonal regeneration,
after intracerebral or intraspinal grafting (277,294). Fetal
remyelination of demyelinated axons, neuroplasticity,
brain tissue, grafted into the CNS of neonatal and adult
neuromodulation, neurogenesis, angiogenesis, anti-inflam-
animals, has been shown to survive and differentiate (318).
matory response, reducing scar and cavity formation,
In brain tissue grafts consisting of undifferentiated ma-
and/or strong phagocytic activity (107,196,203,233,292,
trix cells and few neuroblasts, good development was
293,311,341,388,397,398,419). The cellular composition
observed both in the lateral ventricle and inside the pa-
of the olfactory tissue and the evidence that equivalent
renchyma, 30 and 110 days after transplant. They differ-
cell types exist in both rodent and human olfactory mu-
entiated into organotypical and histotypical structures
cosa suggest that it is potentially a rich source of autolo-
and cells similar to those formed in normal develop-
gous cells for transplant-mediated repair of the CNS
ment. Nerve and glial cells of these transplants were
(224). Selected relevant studies are listed in Table 2.
well differentiated and tightly connected with the sur-
rounding nervous tissue of the host (7). It has been con-vincingly shown that grafting of fetal/embryonic brain
Schwann cells (SCs) play a pivotal role in the mainte-
cells/tissue into the brain and/or spinal cord is a useful
nance and regeneration of the axons in the peripheral
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Table 1. Selected Literature in Preclinical Therapeutic Application of Fetal/Embryonic Brain Cells/Tissue Spinal cord trauma
In vivo magnetic resonance imaging of fetal cat neural tissue trans-plants in the adult cat spinal cord
Monoaminergic reinnervation of the transected spinal cord by ho-mologous fetal brain grafts
Fetal locus coeruleus transplanted into the transected spinal cord ofthe adult rat: Some observations and implications
Visual deficits
Fetal brain tissue transplants reduce visual deficits in adult rats withbilateral lesions of the occipital cortex
Peripheral nerve injury
Viability, growth, and maturation of fetal brain and spinal cord inthe sciatic nerve of adult rat
Parkinson’s disease
Brain grafts reduce motor abnormalities produced by destruction ofnigrostriatal dopamine system
Cognitive deficits
Fetal brain transplant: Reduction of cognitive deficits in rats withfrontal cortex lesions
Fetal brain transplants induce recuperation of taste aversion lear-ning
Brain trauma
Fetal frontal cortex transplanted to injured motor/sensory cortex ofadult rats: Reciprocal connections with host thalamus demonstratedwith WGA-HRP
The effects of intrahippocampal raphe and/or septal grafts in ratswith fimbria-fornix lesions depend on the origin of the grafted tis-sue and the behavioural task used
Organization of host afferents to cerebellar grafts implanted intokainate lesioned cerebellum in adult rats
Hypoxic hypoxia
Transplantation of embryonic brain tissue into the brain of adultrats after hypoxic hypoxia
Neurotrophin-mediated neuroprotection by solid fetal telencephalicgraft in middle cerebral artery occlusion: A preventive approach
Transplantation of human fetal brain cells into ischemic lesions of
nervous system (PNS) due to their ability to dedifferenti-
SC migration and myelination is mediated by interac-
ate, migrate, proliferate, express growth-promoting fac-
tions between sets of extracellular matrix molecules with
tors, and myelinate-regenerating axons (197). Further,
cell surface preoteins, genetic engineering of SCs to al-
SCs have been shown to form myelin after transplanta-
ter aspects of these interactions is a possible way for-
tion into the demyelinated CNS. They can remyelinate
ward. Efforts are, therefore, focused on enhancing their
spinal cord lesions after experimental demyelination,
migration and functional integration into the lesioned
leading in some cases to functional recovery in rodent
CNS. In addition, efforts are under way to use these
and primate models (154). However, SCs do not nor-
cells as tissue engineer seeds and gene delivery vehicles
mally enter the CNS, and migration of SCs transplanted
for an array of molecules with repair potential (33,282).
into the CNS white matter is inhibited by astrocytes. As
The SCs ability to promote restorative efforts has led to
Table 2. Selected Literature in Preclinical Therapeutic Application of Olfactory Ensheathing/Precursor Cells Spinal cord trauma
Regeneration into the spinal cord of transected dorsal root axons ispromoted by ensheathing glia transplants
Ensheathing glia transplants promote dorsal root regeneration andspinal reflex restitution after multiple lumbar rhizotomy
Spinal implants of olfactory ensheathing cells promote axon regen-eration and bladder activity after bilateral lumbosacral dorsal rhi-zotomy in the adult rat
Repair of adult rat corticospinal tract by transplants of olfactoryensheathing cells
Phrenic rehabilitation and diaphragm recovery after cervical injuryand transplantation of olfactory ensheathing cells
Xenotransplantation of transgenic pig olfactory ensheathing cellspromotes axonal regeneration in rat spinal cord
Functional repair of the corticospinal tract by delayed transplanta-tion of olfactory ensheathing cells in adult rats
Protection of corticospinal tract neurons after dorsal spinal cordtransection and engraftment of olfactory ensheathing cells
Long-distance axonal regeneration in the transected adult rat spinalcord is promoted by olfactory ensheathing glia transplants
Functional recovery of paraplegic rats and motor axon regenerationin their spinal cords by olfactory ensheathing glia
Transplantation of nasal olfactory tissue promotes partial recoveryin paraplegic adult rats
Olfactory ensheathing cells promote locomotor recovery after de-layed transplantation into transected spinal cord
Olfactory ensheathing glias transplant improves axonal regenera-tion and functional recovery in spinal cord contusion injury
Delayed transplantation of olfactory ensheathing glia promotessparing/regeneration of supraspinal axons in the contused adult ratspinal cord
Regeneration of adult rat corticospinal axons induced by trans-planted olfactory ensheathing cells
Effects of ensheathing cells transplanted into photochemically dam-aged spinal cord
Increased expression of cyclo-oxygenase 2 and vascular endothelialgrowth factor in lesioned spinal cord by transplanted olfactory en-sheathing cells
Schwann cell-like myelination following transplantation of an ol-
factory bulb-ensheathing cell line into areas of demyelination in theadult CNS
Transplanted olfactory ensheathing cells remyelinate and enhanceaxonal conduction in the demyelinated dorsal columns of the ratspinal cord
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Table 2. Continued
Identification of a human olfactory ensheathing cell that can effecttransplant-mediated remyelination of demyelinated CNS axons
Superparamagnetic iron oxide-labeled Schwann cells and olfactoryensheathing cells can be traced in vivo by magnetic resonance im-aging and retain functional properties after transplantation into theCNS
Remyelination of the nonhuman primate spinal cord by transplanta-tion of H-transferase transgenic adult pig olfactory ensheathingcells
Molecular reconstruction of nodes of Ranvier after remyelinationby transplanted olfactory ensheathing cells in the demyelinated spi-nal cord
Olfactory bulb ensheathing cells enhance peripheral nerve regen-eration
Effect of olfactory ensheathing cells transplantation on protectingspinal cord and neurons after peripheral nerve injury
Transplantation of olfactory ensheathing cells stimulates the collat-eral sprouting from axotomized adult rat facial motoneurons
Transplantation of olfactory mucosa minimizes axonal branchingand promotes the recovery of vibrissae motor performance afterfacial nerve repair in rats
Optic nerve injury
Transplanted olfactory ensheathing cells promote regeneration ofcut adult rat optic nerve axons
Glaucoma
Transplanted olfactory ensheathing cells incorporated into the opticnerve head ensheathe retinal ganglion cell axons: Possible rele-vance to glaucoma
Parkinson’s disease
Olfactory ensheathing cell transplantation restores functional defi-cits in rat model of Parkinson’s disease: A cotransplantation ap-proach with fetal ventral mesencephalic cells
Amyotrophic lateral sclerosis
Adult olfactory bulb neural precursor cell grafts provide temporaryprotection from motor neuron degeneration, improve motor func-tion, and extend survival in amyotrophic lateral sclerosis mice
Cognitive dysfunction
Long-term functional restoration by neural progenitor cell trans-plantation in rat model of cognitive dysfunction: Eo-transplantationwith olfactory ensheathing cells for neurotrophic factor support
A long term observation of olfactory ensheathing cells transplanta-tion to repair white matter and functional recovery in a focal ische-mia model in rat
an increasing interest in using SC grafts for repair of
and unusual antigenic phenotype. Evidence from post-
mortem analysis implicates the involvement of microg-lia in the neurodegenerative process of several neurode-
generative diseases, including AD and PD. Grafting of
Remyelination by transplantation of myelin-forming
activated microglia into the lesioned spinal cord may
cells is possible in animal models; evidence suggests
promote hind limb motor function recovery in rats and
that both a precursor-type oligodendrocyte as well as an
reduce the size of the liquefaction necrosis area (228,
oligodendrocyte that previously formed a myelin sheath
is able to remyelinate the CNS (126,386). Oligodendro-cytes or precursor cells are much more invasive and
have been shown to migrate from the implantation site
Monocytes/macrophages play an integral role in the
to the lesion over a distance of several millimeters
inflammatory process and angiogenesis as well as acting
(72,387). Indeed, a number of studies have demonstrated
as defense mechanisms by exerting microbiocidal and
that transplanted oligodendrocytes survive in the host
immunomodulatory activity. The recruited monocytes/
brain, migrate out of the graft, and synthesize myelin.
macrophages are capable of regulating angiogenesis in
These cells, therefore, have potential for myelin repair
ischemic tissue, tumors, and chronic inflammation. In
after experimental demyelination and in human diseases,
terms of neovascularization followed by tissue regenera-
such as multiple sclerosis, though several findings sug-
tion, monocytes/macrophages should be highly attrac-
gest that OECs and SCs might be more effective than
tive for cell-based therapy compared to other cells due
oligodendrocytes induced from isolated CNS tissue (87,
to their considerable advantages: nononcogenic, nonte-
ratogenic, multiple secretory functions including proan-giogenic and growth factors and their straightforward
cell harvesting procedure (348). Increasing the presence
Cultured astrocytes have been reported to survive and
of activated macrophage/microglial cells at a SCI site
migrate following transplantation. Studies have indi-
can provide an environment beneficial to the promotion
cated that there are differences in the ability of immature
of regeneration of sensory axons, possibly by the release
and mature astrocytes to facilitate plastic changes in the
of cytokines and interaction with other nonneuronal cells
adult brain. Immature astrocytes can synthesize trophic
in the immediate vicinity (117,305).
factors to support neuronal survival, produce a permis-sive environment for neurite extension, and reduce scar
formation. In contrast, mature astrocytes produce a non-
Purkinje cells have a therapeutic value for the re-
permissive environment for axon growth and increase
placement and reconstruction of a defective cerebellar
scar formation. Purified astrocytes were capable of facil-
circuitry in heredodegenerative ataxia. Insignificant
itating behavioral recovery from frontal cortex ablation,
amelioration of motor skills was found in mice after
demyelinating lesions in spinal cord, and kainic acid
solid cerebellar tissue transplantation, while the cell sus-
(KA) lesions of the striatum (10,104,174,240,392). Im-
pension application had no effect (344,357,358).
planted cultured immature astrocytes can stimulate axo-nal regeneration after injury of the postcommissural for-
nix tract in the adult rat brain (406). In addition,
Tanycytes (TAs) are the specialized ependymal glia
behavior alleviation after astrocyte transplantation was
in the CNS, which are located mostly in the ventral wall
shown in rat models of memory deficit induced by
of the third ventricle and median eminence (ME). They
alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionic
closely interact with local cerebrospinal fluid, blood, and
acid lesions, which is independent of cholinergic recov-
neurons. TA is a common component of the brain bar-
ery. The cultured astrocytes may exert their effects over
rier system, the brain–cerebrospinal fluid (CSF) neuro-
a short time period (less than 2 weeks) around the lesion
humoral circuit, and the immune–neuroendocrine net-
site. They can alter the microenvironment and as a result
work. Recent data indicate that TAs transplanted into
less scar tissue was formed followed by less of a barrier
the adult rat spinal cord can support the regeneration of
to the regrowth of nerve fibers (43). Furthermore, evi-
lesioned axons and may represent a useful therapeutic
dence indicates that astrocytes at an immature stage of
differentiation are capable of inducing axon growth fromthe adult optic nerve (354).
Intracerebral transplantation of dopaminergic (DAer-
Microglia are the principal immune cells in the CNS
gic) neuron/cells is currently performed as a restorative
and are characterized by a highly specific morphology
therapy for PD (273). The success of cell therapy in PD
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Table 3. Selected Literature in Preclinical Therapeutic Application of Schwann Cells Spinal cord trauma
Reinnervation of peripheral nerve segments implanted into thehemisected spinal cord estimated by transgenic mice
Schwann cells induce sprouting in motor and sensory axons in theadult rat spinal cord
Influences of the glial environment on the elongation of axons after
injury: Transplantation studies in adult rodents
Axonal regeneration into Schwann cell-seeded guidance channelsgrafted into transected adult rat spinal cord
Reconstruction of the contused cat spinal cord by the delayed nervegraft technique and cultured peripheral non-neuronal cells
Syngeneic grafting of adult rat DRG-derived Schwann cells to theinjured spinal cord
Transplantation of purified populations of Schwann cells into le-sioned adult rat spinal cord
Chronic regenerative changes in the spinal cord after cord compres-sion injury in rats
Remyelination by cells introduced into a stable demyelinating le-sion in the central nervous system
Transplantation of rat Schwann cells grown in tissue culture intothe mouse spinal cord
The use of cultured autologous Schwann cells to remyelinate areas
of persistent demyelination in the central nervous system
Schwann cell remyelination of CNS axons following injection ofcultures of CNS cells into areas of persistent demyelination
Optic nerve injury
Schwann cells and the regrowth of axons in the mammalian CNS:A review of transplantation studies in the rat visual system
Monocular deprivation
Schwann cells transplanted in the lateral ventricles prevent thefunctional and anatomical effects of monocular deprivation in therat
Parkinson’s disease
Cografts of adrenal medulla with peripheral nerve enhance the sur-vivability of transplanted adrenal chromaffin cells and recovery ofthe host nigrostriatal dopaminergic system in MPTP-treated youngadult mice
Peripheral nerve-dopamine neuron co-grafts in MPTP-treated mon-keys: Augmentation of tyrosine hydroxylase-positive fiber stainingand dopamine content in host systems
Therapeutic study of autologous Schwann cells’ bridge graft into
Brain trauma
Transplants of Schwann cell cultures promote axonal regenerationin the adult mammalian brain
Reconstruction of transected postcommissural fornix in adult rat bySchwann cell suspension grafts
Schwann cells transplantation promoted and the repair of brainstem injury in rats
greatly relies on the discovery of an abundant source of
cells capable of DAergic function in the brain. DAergic
ES cells, in particular, possess a nearly unlimited
neuron/precursor cells derived from human embryonic
self-renewal capacity and developmental potential to
stem (hES) cells, human-induced pluripotent stem (hiPS)
differentiate into virtually any cell type of an organism.
cells, neural stem/progenitor cells, human mesenchymal
They can efficiently differentiate into neural precursors,
stem cells, and skin-derived stem cells could be increas-
which can further generate functional neurons, astro-
ingly considered as a pivotal choice for transplant (50,
cytes, and oligodendrocytes (102,229,231). These cells
188,227,269,372). It is likely that cell replacement in
also have the beneficial properties of secreting neuro-
future will focus on not only ameliorating symptoms of
trophic and neural growth factors (272). Along with di-
the disease but also try to slow down the progression of
rected differentiation, other current efforts are aimed at
the disease by either neuroprotection or restoration of a
efficient enrichment, avoidance of immune rejection,
favorable microenvironment in the midbrain (319).
demonstration of functional integration, genetic modifi-cation to regulate neurotransmitter and factor release,
and directed axon growth with these cells (125).
Transplantation of predifferentiated GABAergic neu-
rons significantly induces recovery of sensorimotor func-
tion in brain injury (25). A deficiency of GABAergic
Neural stem/progenitor cells (NSPCs) are present
neurons in the neocortex leads to the dysregulation of
during embryonic development and in certain regions of
cortical neuronal circuits, but this can be overcome by
the adult CNS (264). Mobilizing adult NSCs to promote
cell transplantation. Ventral neural stem cells transfected
repair of injured or diseased parts of the CNS is a prom-
with neurogenin 1 (Ngn1) are integrated as GABAergic
ising approach (3). NSPCs in the adult CNS are capable
neurons within a few days of transplantation into the
of generating new neurons, astrocytes, and oligodendro-
adult mouse neocortex, and the transplantation of com-
cytes (381). Intraventricular transplantation of neural
mitted neuronal progenitor cells has been demonstrated
spheres attenuated brain inflammation in acute and
to be an effective method for brain repair ( 268). In addi-
chronic experimental autoimmune encephalomyelitis
tion, transplants of neuronal cells bioengineered to syn-
(EAE), reduced the clinical severity of disease, and re-
thesize GABA may alleviate chronic neuropathic pain
duced demyelination and axonal pathology. Intravenous
(90). Fetal GABAergic neurons transplanted into the SN
(IV) NSPCs injection also inhibited EAE and reduced
might be an effective means of permanently blocking
CNS inflammation and tissue injury (27). A recent study
seizure generalization in kindling epilepsy and probably
showed that adult NSCs transplanted at sites of injury
also other types of epilepsy (101,234).
can differentiate into vascular cells (endothelial cells andvascular smooth muscle cells) for vasculogenesis (152).
Transplantation of NSCs or their derivatives into a host
Transplanted cholinergic neurons may reinnervate the
brain and the proliferation and differentiation of endoge-
host hippocampus, although this reinnervation appears
nous stem cells by pharmacological manipulations are
to be different from that seen in the intact hippocampal
promising treatments for many neurodegenerative dis-
formation (9). Intraretrosplenial cortical grafts of cholin-
eases and brain injuries, such as PD, brain ischemia, and
ergic neurons can become functionally incorporated
with the host neural circuitry, and the activity of the
implanted cholinergic neurons can be modulated by thehost brain (216) and it can rectify spatial memory defi-
Bone marrow stromal (also called “stem”) cells
cits produced by the loss of intrinsic cholinergic afferents
(BMSCs) can be easily amplified in vitro and their trans-
from the medial septal nucleus (217). Reconstruction of
differentiation into neural cells has been claimed in vitro
the septohippocampal pathways by axons extending
and in vivo (63,82,163,171). The possible mechanisms
from embryonic cholinergic neuroblasts grafted into the
responsible for the beneficial outcome observed after
neuron-depleted septum has been confirmed in the neo-
BMSC transplantation into neurodegenerating tissues in-
natal rat (198). Intrahippocampal septal grafts are able
clude cell replacement, trophic factor delivery, immuno-
to reinnervate the hippocampal formation and ameliorate
modulation, and anti-inflammatory, neuroprotection, and
spatial learning and memory deficits, which are associ-
angiogenesis (171,371,379). Transplantation of BMSCs
ated with anatomical and functional incorporation into
may have a therapeutic role after SCI (64). Adult BMSCs
the circuitry of the host hippocampal formation. Auto-
administered intravenously have been shown to migrate
transplantation of peripheral cholinergic neurons into the
into the brain and improve neurological outcome in rats
cerebral cortex displayed amelioration of abnormal be-
with traumatic brain injury (236). In parallel, data of
intracerebral transplantation suggest that bone marrow
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Table 4. Animal Model of CNS Diseases Treated Using Neural Stem/ Progenitor Cells
could potentially be used to induce plasticity in ischemic
MSC therefore could be a viable alternative to human
brain. Additionally, cotransplantation of BMSCs with
ES cells or NSCs for transplantation therapy of CNS
ES cell-derived graft cells may be useful for preventing
trauma and neurodegenerative diseases (235) (Table 6).
the development of ES cell-derived tumors (253). Re-
sults of this field are summarized in Table 5.
UCB is a rich source of stem cells with great prolifer-
Umbilical Cord Mesenchymal Stromal Cells
ative potential, besides the bone marrow and peripheral
Mesenchymal stromal cells (MSCs) have now been
blood; it has the advantage of being an easily accessible
isolated from most tissues, including the umbilical cord
stem cell source and is less immunogenic compared to
(UC) and UC blood (UCB; see below). UC and UCB
other sources for stem cells (334). There are at least
MSCs are more primitive than those isolated from other
three kinds of stem cells in UCB: hematopoietic, mesen-
tissue sources and do not express the major histocompat-
chymal, and embryonic-like stem cells, which are capa-
ibility complex (MHC) class II human leukocyte anti-
ble of differentiating across tissue lineage boundaries
gen-D-related (HLA-DR) antigens. Studies have shown
into neural, cardiac, epithelial, hepatocytic, and dermal
that UC MSCs are still viable and are not rejected 4
tissue both in vitro and in vivo (132,325,383). Increasing
months after transplantation as xenografts, without the
evidence suggests that MSCs from UCB are present
need for immune suppression, suggesting that they are a
within a wide range of tissues and its therapeutic poten-
favorable cell source for transplantation (422). UC in-
tial extends beyond the hematopoietic component (34).
cluding arteries (UCA), veins (UCV), and Wharton’s
The expanding population of NSPCs can be selected
jelly (UCWJ) is a convenient, efficient source of MSCs
from the human cord blood nonhematopoietic (CD34-
that can be expanded easily in vitro for numerous clini-
negative) mononuclear fraction (49). UCB can be a po-
cal applications for the treatment of nonhematopoietic
tential source for autologous or allogeneic monocytes/
diseases, and in studies of tissue regeneration and immu-
macrophages. UCB monocytes should be considered as
nosuppression (119,159). UC MSCs have proven to be
a primary candidate owing to their easy isolation, low
efficacious in reducing lesion sizes and enhancing be-
immune rejection, and multiple characteristic advan-
havioral recovery in animal models of ischemic and
tages such as their anti-inflammatory properties by
traumatic CNS injury. Recent findings also suggest that
virtue of their unique immune and inflammatory imma-
neurons derived from UC-MSC could alleviate move-
turity, and their proangiogenic ability (333). The thera-
ment disorders in hemiparkinsonian animal models. UC-
peutic potential of UCB cells may be attributed to the
Table 5. Animal Model of CNS Diseases Treated Using Bone Marrow Stromal Cells
Experimental autoimmune encephalomyelitis (EAE)
inherent ability of stem cell populations to replace dam-
1995 based on favorable results in animal models in-
aged tissues. Alternatively, various cell types within the
cluding EAE (47). Recent studies show that transplanta-
graft may promote neural repair by delivering neural
tion of HSCs from bone marrow is an effective strategy
protection and secretion of neurotrophic factors (284,
for SCI after directly transplanting cells into the cord
334). In addition, evidence suggests that delivery of cir-
1 week after injury (185), with a similar potential in
culating CD34+ human UCB cells can produce func-
comparison with marrow stromal cells (180). An in-
tional recovery in an animal stroke model with concur-rent angiogenesis and neurogenesis leading to somerestoration of cortical tissue (295). UCB cells have been
Table 7. Animal Model of CNS Diseases Treated Using
used in preclinical models of brain injury and neurode-
generative diseases, directed to differentiate into neural
phenotypes, and have been related to functional recov-
ery after engraftment in CNS lesion models (Table 7).
Hematopoietic stem cell transplantation (HSCT) was
proposed as a treatment for multiple sclerosis (MS) in
Table 6. Animal Model of CNS Diseases Treated Using
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
creasing number of studies provide evidence that hema-
myelinated endogenous host axons, recruited endoge-
topoietic stem cells, either after stimulation of endoge-
nous SCs into the injured cord, formation of a bridge
nous stem cell pools or after exogenous hematopoietic
across the lesion site, increased size of the spared tissue
stem cell use, improve functional outcome after ische-
rim, myelinated spared axons within the tissue rim, re-
mic brain lesions. Various underlying mechanisms such
duced reactive gliosis, and an environment that was
as transdifferentiation into neural lineages, neuroprotec-
highly conducive to axonal growth (35). In addition,
tion through trophic support, and cell fusion have been
SKPs transplanted into PD model rats sufficiently differ-
deciphered (130). Furthermore, intracerebral peripheral
entiated into dopamine neuron-like cells, and partially
blood hematopoietic stem cell (CD34+) implantation in-
but significantly corrected their behavior. The generated
duces neuroplasticity by enhancing β1 integrin-mediated
DA neuron-like cells are expected to serve as donor cells
angiogenesis in chronic cerebral ischemia with signifi-
for neuronal repair for PD (188). Thus, this cell line has
cantly increased modulation of neurotrophic factor ex-
been identified as novel, accessible, and a potentially
pression in the ischemic hemisphere (352).
autologous source for future nervous system repair(35,192). Adipose-Derived Adult Stem/Precursor Cells
Adipose tissue is an abundant, accessible, and replen-
The retinal pigment epithelium consists of a unicellu-
ishable source of adult stem cells that can be isolated
lar layer of neuroepithelial cells, retinal pigment epithe-
from liposuction waste tissue by collagenase digestion
lial (RPE) cells, which are essential for the maintenance
and differential centrifugation (118). These adipose-
of the normal function of the retina (139). Cultured hu-
derived adult stem (ADAS) cells, which exhibit charac-
man RPE cells have the capacity to synthesize neuro-
teristics of multipotent adult stem cells, similar to those
trophins, including NGF, brain-derived growth factor
of MSCs, are multipotent, differentiating along the adi-
(BDNF), glial cell-derived neurotrophic factor (GDNF),
pocyte, chondrocyte, myocyte, neuronal, and osteoblast
and neurotrophin-3 (NT-3) (158,262). Studies have
lineages (124). ADAS cells have potential applications
shown that, as an alternative cell source, RPE cells pos-
for the repair and regeneration of acute and chronically
sess DAergic replacement properties with neurotrophic
damaged tissues (329). As an alternative stem cell
support on primary cultures of rat striatal (enkephaliner-
source for CNS therapies, ADAS cells labeled with su-
gic) and mesencephalic (DAergic) neurons, and there-
perparamagnetic iron oxide have been shown using MRI
fore could exert a positive effect in parkinsonian animals
to successfully transplant in vivo in unilateral middle
by intrastriatal transplantation (247,257,365,366). RPE
cerebral artery occluded (MCAo) mice (320). The study
cells can be transduced with high efficiency using an
of Ryu and colleagues indicate that improvement in neu-
adenoviral vector, making them promising vehicles for
rological function by the transplantation of ADAS in
local delivery of therapeutic proteins for the treatment
dogs with SCI may be partially due to the neural differ-
of neurodegenerative diseases in a combined cell and
entiation of the implanted stem cells (326). Furthermore,
the transplantation of ADAS can promote the formationof a more robust nerve in rats with a sciatic nerve defect
and produce a decrease in muscle atrophy (336).
Human amniotic epithelial cells (AEC) do not ex-
press the HLA-A, -B, -C, or -DR antigens on their sur-
face, which suggests no acute rejection in transplanta-
Skin-derived precursors (SKPs) are a self-renewing,
tion (6). The human amnion membrane serves as a
multipotent precursor that are generated during embryo-
bridge for axonal regeneration in vitro and in vivo; cells
genesis and persist into adulthood in the dermis, share
isolated from the amniotic membrane can differentiate
characteristics with embryonic neural crest stem cells,
into all three germ layers, have low immunogenicity and
including their ability to differentiate into neural crest-
anti-inflammatory function (76,417). Given their multi-
derived cell types such as peripheral neurons, SCs,
potent differentiation ability, capability of synthesizing
astrocytes, and endothelium (26,149). After transplanta-
catecholamines including DA, and neurotrophic and
tion, the cells yield healthy cells that migrate to the le-
neuroprotection effect, there is accumulating evidence
sion site, and then differentiate mainly into cells ex-
that suggests that AECs have therapeutic potential for
pressing glia and neuronal markers (122). Recent
multiple CNS disorders, such as PD, mucopolysacchari-
evidence indicates that transplantation of SKP-derived
dosis, SCI, stroke, brain trauma, etc. (Table 8).
SCs represent a viable alternative strategy for repairing
the injured spinal cord, with the neuroanatomical neur-orestorative findings including good survival within the
Endometrial cells supplied as a form of menstrual
injured spinal cord, reduced size of the contusion cavity,
blood–tissue mixture can be used for cell-based restor-
Table 8. Selected Literatures in Preclinical Therapeutic Application of Amniotic Epithelial Cells Parkinson’s disease
Human amniotic epithelial cells produce dopamine and surviveafter implantation into the striatum of a rat model of Parkinson’sdisease: A potential source of donor for transplantation therapy
Implantation of human amniotic epithelial cells prevents the degen-eration of nigral dopamine neurons in rats with 6-hydroxydopaminelesions
Transplantation of human amniotic cells exerts neuroprotection inMPTP-induced Parkinson disease mice
Mucopolysaccharidosis
Engraftment of genetically engineered amniotic epithelial cells cor-
rects lysosomal storage in multiple areas of the brain in mucopoly-saccharidosis type VII mice
Brain ischemia
Amniotic epithelial cells transform into neuron-like cells in the is-chemic brain
Amniotic fluid derived stem cells ameliorate focal cerebral isch-aemia-reperfusion injury induced behavioural deficits in mice
Human amniotic epithelial cells ameliorate behavioral dysfunctionand reduce infarct size in the rat middle cerebral artery occlusionmodel
Spinal cord trauma
Role of human amniotic epithelial cell transplantation in spinal cordinjury repair research
Transplantation of human amniotic epithelial cells improves hind-limb function in rats with spinal cord injury
Peripheral nerve injury
Bridging rat sciatic nerve defects with the composite nerve-muscleautografts wrapped with human amnion matrix membrane
Brain injury
Treatment of traumatic brain injury in rats with transplantation ofhuman amniotic cells
ative therapy in muscular dystrophy (380); subsequent
evidence shows that populations of stromal stem cellsderived from menstrual blood are multipotent, being
Transplanted testis-derived Sertoli cells, which create
able to differentiate into chondrogenic, adipogenic, os-
a localized immune “privileged” site, possess a modula-
teogenic, neurogenic, and cardiogenic cell lineages (290).
tory function on graft rejection and survival and act as
The cultured menstrual blood express embryonic like-
a viable graft source for facilitating the use of xenotrans-
stem cell phenotypic markers [Octamer-4 (Oct4), stage-
plantation for diabetes, PD, Huntington’s disease, and
specific embryonic antigen (SSEA), Nanog], and when
other neurodegenerative diseases (41,331,332). In addi-
grown in appropriate conditioned media, express neu-
tion to producing immunoprotective factors, Sertoli cells
ronal phenotypic markers [nestin, microtubule-associ-
also secrete growth and trophic factors that appear to
ated protein 2 (MAP2)] (40). Transplantation of men-
enhance the posttransplantation viability of isolated cells
strual blood-derived stem cells, either intracerebrally or
and, likewise, the postthaw viability of isolated, cryopre-
intravenously and without immunosuppression, signifi-
served cells (52). Sertoli cells grafted into adult rat
cantly reduces behavioral and histological impairments
brains ameliorated behavioral deficits and enhanced
DAergic neuronal survival and outgrowth (331). Cotrans-
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
planting of Sertoli cells may be useful as a combination
younger, but not in older patients (108). Furthermore,
therapy in CNS lesions, a strategy that could enhance
pathologic findings suggest that grafts of fetal mesence-
the recovery benefits associated with transplantation and
phalic DA neurons could survive long term with or with-
decrease the need for, and the risks associated with,
out α-synuclein-positive Lewy bodies (184,205,259).
long-term systemic immunosuppression (399). Further,
Bachoud-Le´vi and coworkers indicated motor and
recent research has shown that implantation of a Sertoli
cognitive recovery in patients with Huntington’s disease
cell-enriched preparation has a significant neuroprotec-
after neural transplantation (17), which continued during
tive benefit to vulnerable motor neurons in a superoxide
long-term follow-up (16). The therapeutic value of hu-
dismutase 1 (SOD1) transgenic mouse model of amyo-
man striatal neuroblasts in Huntington’s disease was
trophic lateral sclerosis (ALS) (136).
identified by Gallina and colleagues (112). Human Neural Stem/Progenitor Cells
Induced pluripotent stem (iPS) cells are derived from
somatic cells by ectopic expression of a few transcrip-
The clinical regimes of intracranial implant of human
tion factors. iPS cells appear to be able to self-renew
neuronal cells in stroke patients have proven safe and
indefinitely and to differentiate into all types of cells
feasible, though there is a lack of evidence for a signifi-
in the body, and are almost identical to ES cells. The
cant benefit in motor function (181,182). Data suggest
generation of patient-derived pluripotent cells applicable
that cell therapy is a safe method and can be effectively
to autologous cell-based therapies has the potential to
used for stroke (308), acute brain injury (346,347), and
revolutionize medicine (60). Since the first report from
cerebral palsy (345). In addition, neurological function
Takahashi and Yamanaka on the reprogramming of
has been restored after autologous neural stem cell trans-
mouse fibroblasts into pluripotent stem cells by defined
plantation in patients with brain trauma (428).
factors in 2006 (370), various new methods have beendeveloped to refine and improve reprogramming tech-
Umbilical Cord Mesenchymal Stem Cells
nology (281). The current demonstration of DAergic dif-ferentiation of human induced pluripotent stem cells
Therapy of UC-MSCs could stabilize the disease
(hiPSCs), replacement of segmental losses of interneur-
course of refractory progressive MS (218).
ons and motorneurons due to gray matter damage andrestoration of auditory spiral ganglion neurons suggest a
Umbilical Cord Blood Mesenchymal Stem Cells
new avenue for highly effective, tumor-free, and im-
Kang and colleagues report that UCB-MSC trans-
mune rejection-free cell therapy for PD, SCI, and hear-
plantation may play a role in the treatment of SCI pa-
ing disturbance in the near future (151,276,323). CLINICAL STUDIES OF CELL-BASED Olfactory Ensheathing Cell and OlfactoryNEURORESTORATOLOGY IN CNS DISEASES (TABLE 9)
Early OEC/olfactory mucosa autograft transplants for
Adrenal Medullary Tissue, Substantia Nigra,
patients with chronic SCI were reported by Huang et al.
in 2003 (143), Rabinovich et al. in 2003 (309), and Lima
Positive findings had initially been observed by
et al. in 2006 (223), and the results were safe, feasible,
Backlund et al. (19) and Lindvall et al. (225) concerning
and positive. Mackay-Sim and colleagues reported au-
the transplantation of autologous adrenal medullary tis-
tologous OEC transplantation for three patients with
sue into the striatum of patients with severe parkinson-
chronic SCI with 3-year follow-up was safe, feasible,
ism. Subsequently, Hitchcock et al. (138), Lindvall and
and one patient showed sensory improvement (243). The
colleagues (226,395), and Madrazo et al. (244) have sep-
therapeutic value of OEC transplantation has been
arately reported that fetal nigral implants might have
shown in chronic SCI, ALS, cerebral palsy, stroke, MS,
provided a modest improvement in motor function and
and other neurodegenerative diseases and traumatic
have clinically valuable improvements in most recipi-
brain insults in 1,255 patients (144,145).
ents within a period of long-term follow-up after trans-plantation into the brain of patients with PD. Freed and
colleagues randomly assigned patients to receive nervecell transplants or sham surgery with double-blind fol-
Data from Arjmand and colleagues suggested that au-
low-up. The result showed that transplanted human em-
tologous SC transplantation is safe for spinal cord in-
bryonic DA neurons survive with clinical benefit in
jured patients but had no beneficial effects (327). Table 9. Selected Recent Articles of Clinical Studies Related to Cell-Based CNS Neurorestoratology
lateral sclerosis, cerebral palsy,multiple sclerosis, stoke, etc.
by Farge et al. (98), Fassas et al. (99), Portaccio et al. (304), and Saccardi et al. (328) as well as in studies for
Knoller et al. reported that autologous macrophages
malignant or severe MS (54,177,246).
Nonmyeloablative autologous haemopoietic stem cell
Bone Marrow Stromal Cell/Hematopoietic Stem
transplantation in patients with relapsing-remitting MS
reverses neurological deficits (48). Also autologous pe-ripheral blood stem cell transplantation has promoted
Research of Appel et al. indicated that peripheral
obvious neurologic improvement for patients with poly-
cells derived from donor hematopoietic stem cells were
neuropathy, organomegaly, endocrinopathy, M-protein,
able to enter the human CNS primarily at sites of moto-
and skin changes syndrome (190,191).
neuron pathology and engrafted as immunomodulatorycells, but they did not provide benefit in sporadic ALS
SUMMARY AND PROSPECTS
patients (12). On the contrary, autologous anti-humanCD133+ mononuclear cell transplantation in the motor
When entering the 21st century, numerous centers
cortex delays ALS progression and improves quality of
have globally started clinical trials or experimental treat-
life (251). Furthermore, many studies showed that this
ments to investigate the utilization of cells, such as neu-
kind of cell therapy was feasible, safe, and effective for
rons, OECs, bone marrow-derived cells, NSPCs, SCs,
ALS (78), stroke (21,364), chronic SCI patients (61,77,
etc., for intractable CNS diseases. Despite their diversity
266,287,368), and there was also improvement in the
in number, clinical status of subjects, route of cell ad-
acute and subacute phase of chronic SCI (416). Different
ministration, and criteria to evaluate efficacy, the main
routes of cell transplantation such as by direct injection
conclusion drawn from these clinical studies was that
into spinal cord, intravenous and intrathecal injection
such therapies were safe, feasible, and had some neuro-
have proven to be equally effective in SCI (116) and
logical functional improvement or restorative effect that
traumatic brain injury (424). Evidence shows that autol-
improved the patient’s quality of life to a varying extent.
ogous hematopoietic stem cell transplantation cannot be
These achievements had already answered YES or NO
deemed a curative treatment but instead may give rise
to the question of whether the degeneration and damage
to prolonged stabilization or change the aggressive
in the CNS could be functionally restored.
course of diseases (330). Similar results were reported
But from the cellular biology viewpoint, there are
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
jury-scientific challenges for the unknown future. Ups. J.
several unanswered questions for cell transplantation:
what kind of cells would be the best ideal source, the
9. Anderson, K. J.; Gibbs, R. B.; Salvaterra, P. M.; Cotman,
best therapeutic time window, the most suitable selec-
C. W. Ultrastructural characterization of identified cho-
tion for patients and diseases of different kinds, and the
linergic neurons transplanted to the hippocampal forma-
optimal route. Consequently, emphasis should be placed
tion of the rat. J. Comp. Neurol. 249(2):279–292; 1986.
10. Andersson, C.; Tytell, M.; Brunso-Bechtold, J. Trans-
on solving these questions and evaluating the efficacy
plantation of cultured type 1 astrocyte cell suspensions
of each particular treatment modality in detail.
into young, adult and aged rat cortex: Cell migration and
On the other hand, from a clinical neurorestoratology
survival. Int. J. Dev. Neurosci. 11(5):555–568; 1993.
viewpoint, the current treatment results are far from an
11. Andres, R. H.; Meyer, M.; Ducray, A. D.; Widmer,
effective cure or the miracle effect, as the majority of
H. R. Restorative neuroscience: Concepts and perspec-tives. Swiss Med. Wkly. 138(11–12):155–172; 2008.
people expect; however, therapeutic strategies to retard
12. Appel, S. H.; Engelhardt, J. I.; Henkel, J. S.; Siklos, L.;
disease progression for neurodegenerative diseases or to
Beers, D. R.; Yen, A. A.; Simpson, E. P.; Luo, Y.;
improve some functions from acquired damages seem to
Carrum, G.; Heslop, H. E.; Brenner, M. K.; Popat, U.
be a more realistic clinical aim compared with expecting
Hematopoietic stem cell transplantation in patients with
a cure or complete recovery in the present or near future.
sporadic amyotrophic lateral sclerosis. Neurology 71(17):1326–1334; 2008.
Patients, scientists, and doctors should value highly the
13. Armengol, J. A.; Sotelo, C.; Angaut, P.; Alvarado-
patients’ achievements from effective treatment strate-
Mallart, R. M. Organization of host afferents to cerebel-
gies that are currently still thought by some to not be
lar grafts implanted into kainate lesioned cerebellum in
adult rats. Eur. J. Neurosci. 1(1):75–93; 1989.
So far, the pleasurable reality is that landmark ad-
14. Arnhold, S.; Semkova, I.; Andressen, C.; Lenartz, D.;
Meissner, G.; Sturm, V.; Kochanek, S.; Addicks, K.;
vances and the results of preclinical and clinical studies
Schraermeyer, U. Iris pigment epithelial cells: A possible
in neurorestoratolgy have been driving our traditional
cell source for the future treatment of neurodegenerative
concept from the passive reaction to disease to active
diseases. Exp. Neurol. 187(2):410–417; 2004.
attempts to restore the lost functions of the CNS. Now
15. Auerbach, J. M.; Eiden, M. V.; McKay, R. D. Trans-
people are more interested in and pay more attention
planted CNS stem cells form functional synapses in vivo. Eur. J. Neurosci. 12(5):1696–1704; 2000.
to functional neurorestoration rather than the anatomical
Lefaucheur, J. P.; Boisse´, M. F.; Maison, P.; Baudic, S.;Ribeiro, M. J.; Bourdet, C.; Remy, P.; Cesaro, P.;
REFERENCES
Hantraye, P.; Peschanski, M. Effect of fetal neural trans-
1. Agrawal, A. K.; Shukla, S.; Chaturvedi, R. K.; Seth, K.;
plants in patients with Huntington’s disease 6 years after
Srivastava, N.; Ahmad, A.; Seth, P. K. Olfactory en-
surgery: A long-term follow-up study. Lancet Neurol.
sheathing cell transplantation restores functional deficits
in rat model of Parkinson’s disease: A cotransplantation
17. Bachoud-Le´vi, A. C.; Re´my, P.; Nguyen, J. P.;
approach with fetal ventral mesencephalic cells. Neuro-
Brugie`res, P.; Lefaucheur, J. P.; Bourdet, C.; Baudic, S.;
Gaura, V.; Maison, P.; Haddad, B.; Boisse´, M. F.;
2. Aguayo, A. J.; David, S.; Bray, G. M. Influences of the
Grandmougin, T.; Je´ny, R.; Bartolomeo, P.; Dalla Barba,
glial environment on the elongation of axons after injury:
G.; Degos, J. D.; Lisovoski, F.; Ergis, A. M.; Pailhous,
Transplantation studies in adult rodents. J. Exp. Biol. 95:
E.; Cesaro, P.; Hantraye, P.; Peschanski, M. Motor and
cognitive improvements in patients with Huntington’s
3. Ahmed, S. The culture of neural stem cells. J. Cell. Bio-
disease after neural transplantation. Lancet 356(9246):
4. Akerud, P.; Canals, J. M.; Snyder, E. Y.; Arenas, E.
18. Bachstetter, A. D.; Pabon, M. M.; Cole, M. J.; Hudson,
Neuroprotection through delivery of glial cell line-
C. E.; Sanberg, P. R.; Willing, A. E.; Bickford, P. C.;
derived neurotrophic factor by neural stem cells in a
Gemma, C. Peripheral injection of human umbilical cord
mouse model of Parkinson’s disease. J. Neurosci.
blood stimulates neurogenesis in the aged rat brain. BMC
5. Akiyama, Y.; Radtke, C.; Kocsis, J. D. Remyelination of
19. Backlund, E. O.; Granberg, P. O.; Hamberger, B.; Knuts-
the rat spinal cord by transplantation of identified bone
son, E.; Ma˚rtensson, A.; Sedvall, G.; Seiger, A.; Olson,
marrow stromal cells. J. Neurosci. 22(15):6623–6630;
L. Transplantation of adrenal medullary tissue to stria-
tum in parkinsonism. First clinical trials. J. Neurosurg.
6. Akle, C. A.; Adinolfi, M.; Welsh, K. I.; Leibowitz, S.;
McColl, I. Immunogenicity of human amniotic epithelial
20. Bakshi, A.; Barshinger, A. L.; Swanger, S. A.; Madha-
vani, V.; Shumsky, J. S.; Neuhuber, B.; Fischer, I. Lum-
bar puncture delivery of bone marrow stromal cells in
7. Alexandrova, M. A.; Polezhaev, L. V. Transplantation of
spinal cord contusion: A novel method for minimally in-
various regions of embryonic brain tissue into the brain
vasive cell transplantation. J. Neurotrauma 23(1):55–65;
of adult rats. J. Hirnforsch. 25(1):89–98; 1984.
8. Anderberg, L.; Aldskogius, H.; Holtz, A. Spinal cord in-
21. Bang, O. Y.; Lee, J. S.; Lee, P. H.; Lee, G. Autologous
mesenchymal stem cell transplantation in stroke patients.
Schwann cell remyelination of CNS axons following in-
jection of cultures of CNS cells into areas of persistent
22. Bantubungi, K.; Blum, D.; Cuvelier, L.; Wislet-Gende-
demyelination. Neurosci. Lett. 77(1):20–24; 1987.
bien, S.; Rogister, B.; Brouillet, E.; Schiffmann, S. N.
38. Blo¨mer, U.; Naldini, L.; Verma, I. M.; Trono, D.; Gage,
Stem cell factor and mesenchymal and neural stem cell
F. H. Applications of gene therapy to the CNS. Hum.
transplantation in a rat model of Huntington’s disease.
Mol. Cell. Neurosci. 37(3):454–470; 2008.
39. Borlongan, C. V.; Hadman, M.; Sanberg, C. D.; Sanberg,
23. Barami, K.; Hao, H. N.; Lotoczky, G. A.; Diaz, F. G.;
P. R. Central nervous system entry of peripherally in-
Lyman, W. D. Transplantation of human fetal brain cells
jected umbilical cord blood cells is not required for neu-
into ischemic lesions of adult gerbil hippocampus. J.
roprotection in stroke. Stroke 35(10):2385–2389; 2004.
40. Borlongan, C. V.; Kaneko, Y.; Maki, M.; Yu, S.; Ali,
24. Barnett, S. C.; Alexander, C. L.; Iwashita, Y.; Gilson,
M. M.; Allickson, J.; Sanberg, C.; Kuzmin-Nichols, N.;
J. M.; Crowther, J.; Clark, L.; Dunn, L. T.; Papanastas-
Sanberg, P. R. Menstrual blood cells display stem cell-
siou, V.; Kennedy, P. G.; Franklin, R. J. Identification of
like phenotypic markers and exert neuroprotection fol-
a human olfactory ensheathing cell that can effect trans-
lowing transplantation in experimental stroke. Stem Cells
plant-mediated remyelination of demyelinated CNS ax-
ons. Brain 123(Pt. 8):1581–1588; 2000.
41. Borlongan, C. V.; Stahl, C. E.; Cameron, D. F.; Saporta,
25. Becerra, G. D.; Tatko, L. M.; Pak, E. S.; Murashov,
S.; Freeman, T. B.; Cahill, D. W.; Sanberg, P. R. CNS
A. K.; Hoane, M. R. Transplantation of GABAergic neu-
immunological modulation of neural graft rejection and
rons but not astrocytes induces recovery of sensorimotor
survival. Neurol. Res. 18(4):297–304; 1996.
function in the traumatically injured brain. Behav. Brain
42. Bottai, D.; Madaschi, L.; Di Giulio, A. M.; Gorio, A.
Viability-dependent promoting action of adult neural
26. Belicchi, M.; Pisati, F.; Lopa, R.; Porretti, L.; Fortunato,
precursors in spinal cord injury. Mol. Med. 14(9–10):
F.; Sironi, M.; Scalamogna, M.; Parati, E. A.; Bresolin,
N.; Torrente, Y. Human skin-derived stem cells migrate
43. Bradbury, E. J.; Kershaw, T. R.; Marchbanks, R. M.;
throughout forebrain and differentiate into astrocytes
Sinden, J. D. Astrocyte transplants alleviate lesion in-
after injection into adult mouse brain. J. Neurosci. Res.
duced memory deficits independently of cholinergic re-
covery. Neuroscience 65(4):955–972; 1995.
27. Ben-Hur, T. Immunomodulation by neural stem cells. J.
44. Bretzner, F.; Liu, J.; Currie, E.; Roskams, A. J.; Tetzlaff,
Neurol. Sci. 265(1–2):102–104; 2008.
W. Undesired effects of a combinatorial treatment for
28. Ben-Hur, T.; Goldman, S. A. Prospects of cell therapy
spinal cord injury-transplantation of olfactory ensheath-
for disorders of myelin. Ann. NY Acad. Sci. 1142:218–
ing cells and BDNF infusion to the red nucleus. Eur. J.
29. Bermu´dez-Rattoni, F.; Ferna´ndez, J.; Sa´nchez, M. A.;
45. Brunet, J. F.; Redmond, Jr., D. E.; Bloch, J. Primate
Aguilar-Roblero, R.; Drucker-Colı´n, R. Fetal brain trans-
adult brain cell autotransplantation, a pilot study in
plants induce recuperation of taste aversion learning.
asymptomatic MPTP-treated monkeys. Cell Transplant.
30. Bernstein, J. J. Viability, growth, and maturation of fetal
46. Bunge, M. B. Transplantation of purified populations of
brain and spinal cord in the sciatic nerve of adult rat. J.
Schwann cells into lesioned adult rat spinal cord. J. Neu-
Neurosci. Res. 10(4):343–350; 1983.
rol. 242(1 Suppl. 1):S36–S39; 1994.
31. Bernstein, J. J.; Goldberg, W. J. Fetal spinal cord homo-
47. Burt, R. K.; Burns, W.; Hess, A. Bone marrow transplan-
grafts ameliorate the severity of lesion-induced hind limb
tation for multiple sclerosis. Bone Marrow Transplant.
behavioral deficits. Exp. Neurol. 98(3):633–644; 1987.
32. Bernstein-Goral, H.; Bregman, B. S. Spinal cord trans-
48. Burt, R. K.; Loh, Y.; Cohen, B.; Stefoski, D.; Balabanov,
plants support the regeneration of axotomized neurons
R.; Katsamakis, G.; Oyama, Y.; Russell, E. J.; Stern, J.;
after spinal cord lesions at birth: A quantitative double-
Muraro, P.; Rose, J.; Testori, A.; Bucha, J.; Jovanovic,
labeling study. Exp. Neurol. 123(1):118–132; 1993.
B.; Milanetti, F.; Storek, J.; Voltarelli, J. C.; Burns,
33. Bhatheja, K.; Field, J. Schwann cells: Origins and role
W. H. Autologous non-myeloablative haemopoietic stem
in axonal maintenance and regeneration. Int. J. Biochem.
cell transplantation in relapsing-remitting multiple scle-
Cell Biol. 38(12):1995–1999; 2006.
rosis: A phase I/II study. Lancet Neurol. 8(3):244–253;
34. Bieback, K.; Klu¨ter, H. Mesenchymal stromal cells from
umbilical cord blood. Curr. Stem Cell Res. Ther. 2(4):
49. Buzan´ska, L.; Jurga, M.; Doman´ska-Janik, K. Neuronal
differentiation of human umbilical cord blood neural
35. Biernaskie, J.; Sparling, J. S.; Liu, J.; Shannon, C. P.;
stem-like cell line. Neurodegener. Dis. 3(1–2):19–26;
Plemel, J. R.; Xie, Y.; Miller, F. D.; Tetzlaff, W. Skin-
derived precursors generate myelinating Schwann cells
50. Cai, J.; Yang, M.; Poremsky, E.; Kidd, S.; Schneider,
that promote remyelination and functional recovery after
J. S.; Iacovitti, L. Dopaminergic neurons derived from
contusion spinal cord injury. J. Neurosci. 27(36):9545–
human induced pluripotent stem cells survive and inte-
grate into 6-OHDA lesioned rats. Stem Cells Dev. in
36. Blakemore, W. F.; Crang, A. J. The use of cultured au-
tologous Schwann cells to remyelinate areas of persistent
51. Cajal, S. R. Y. Degeneration and regeneration of the ner-
demyelination in the central nervous system. J. Neurol.
vous system (R. M. May, Trans.). London: Oxford Uni-
37. Blakemore, W. F.; Crang, A. J.; Patterson, R. C.
52. Cameron, D. F.; Othberg, A. I.; Borlongan, C. V.;
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Rashed, S.; Anton, A.; Saporta, S.; Sanberg, P. R. Post-
J. H.; Hwang, S. H.; Han, H.; Lee, J. H.; Choe, B. Y.;
thaw viability and functionality of cryopreserved rat fetal
Lee, S. Y.; Kim, H. Y. Intraarterially delivered human
brain cells cocultured with Sertoli cells. Cell Transplant.
umbilical cord blood-derived mesenchymal stem cells in
canine cerebral ischemia. J. Neurosci. Res. 87(16):3554–
53. Cao, Q. L.; Zhang, Y. P.; Howard, R. M.; Walters,
W. M.; Tsoulfas, P.; Whittemore, S. R. Pluripotent stem
66. Cicchetti, F.; Saporta, S.; Hauser, R. A.; Parent, M.;
cells engrafted into the normal or lesioned adult rat spi-
Saint-Pierre, M.; Sanberg, P. R.; Li, X. J.; Parker, J. R.;
nal cord are restricted to a glial lineage. Exp. Neurol.
Chu, Y.; Mufson, E. J.; Kordower, J. H.; Freeman, T. B.
Neural transplants in patients with Huntington’s disease
54. Capello, E.; Saccardi, R.; Murialdo, A.; Gualandi, F.;
undergo disease-like neuronal degeneration. Proc. Natl.
Pagliai, F.; Bacigalupo, A.; Marmont, A.; Uccelli, A.;
Acad. Sci. USA 106(30):12483–12488; 2009.
Inglese, M.; Bruzzi, P.; Sormani, M. P.; Cocco, E.;
67. Collier, T. J.; Elsworth, J. D.; Taylor, J. R.; Sladek, Jr.,
Meucci, G.; Massacesi, L.; Bertolotto, A.; Lugaresi, A.;
J. R.; Roth, R. H.; Redmond, Jr., D. E. Peripheral nerve-
Merelli, E.; Solari, A.; Filippi, M.; Mancardi, G. L.; Ital-
dopamine neuron co-grafts in MPTP-treated monkeys:
ian GITMO-Neuro Intergroup on ASCT for Multiple
Augmentation of tyrosine hydroxylase-positive fiber
Sclerosis. Intense immunosuppression followed by autol-
staining and dopamine content in host systems. Neuro-
ogous stem cell transplantation in severe multiple sclero-
sis. Neurol. Sci. 26(Suppl. 4):S200–S203; 2005.
68. Commissiong, J. W. Fetal locus coeruleus transplanted
55. Chen, J.; Li, Y.; Katakowski, M.; Chen, X.; Wang, L.;
into the transected spinal cord of the adult rat: Some ob-
Lu, D.; Lu, M.; Gautam, S. C.; Chopp, M. Intravenous
servations and implications. Neuroscience 12(3):839–
bone marrow stromal cell therapy reduces apoptosis and
promotes endogenous cell proliferation after stroke in fe-
69. Coronel, M. F.; Musolino, P. L.; Villar, M. J. Selective
male rat. J. Neurosci. Res. 73(6):778–786; 2003.
migration and engraftment of bone marrow mesenchy-
56. Chen, J.; Li, Y.; Wang, L.; Zhang, Z.; Lu, D.; Lu, M.;
mal stem cells in rat lumbar dorsal root ganglia after sci-
Chopp, M. Therapeutic benefit of intravenous adminis-
atic nerve constriction. Neurosci. Lett. 405(1–2):5–9;
tration of bone marrow stromal cells after cerebral ische-
mia in rats. Stroke 32(4):1005–1011; 2001.
70. Corti, S.; Locatelli, F.; Donadoni, C.; Guglieri, M.;
57. Chen, J.; Sanberg, P. R.; Li, Y.; Wang, L.; Lu, M.; Will-
Papadimitriou, D.; Strazzer, S.; Del Bo, R.; Comi, G. P.
ing, A. E.; Sanchez-Ramos, J.; Chopp, M. Intravenous
Wild-type bone marrow cells ameliorate the phenotype
administration of human umbilical cord blood reduces
of SOD1-G93A ALS mice and contribute to CNS, heart
behavioral deficits after stroke in rats. Stroke 32(11):
and skeletal muscle tissues. Brain 127(Pt. 11):2518–
58. Chen, J.; Zhang, Z. G.; Li, Y.; Wang, L.; Xu, Y. X.;
71. Corti, S.; Nizzardo, M.; Nardini, M.; Donadoni, C.;
Gautam, S. C.; Lu, M.; Zhu, Z.; Chopp, M. Intravenous
Salani, S.; Ronchi, D.; Saladino, F.; Bordoni, A.; Fortu-
administration of human bone marrow stromal cells in-
nato, F.; Del Bo, R.; Papadimitriou, D.; Locatelli, F.;
duces angiogenesis in the ischemic boundary zone after
Menozzi, G.; Strazzer, S.; Bresolin, N.; Comi, G. P. Neu-
stroke in rats. Circ. Res. 92:692–699; 2003.
ral stem cell transplantation can ameliorate the pheno-
59. Chen, L.; Huang, H. Neurorestoratology: New concept
type of a mouse model of spinal muscular atrophy. J.
and bridge from bench to bedside. Zhongguo Xiu Fu
Clin. Invest. 118(10):3316–3330; 2008.
Chong Jian Wai Ke Za Zhi 23(3):366–370; 2009.
72. Crang, A. J.; Blakemore, W. F. Remyelination of demye-
60. Chen, L.; Liu, L. Current progress and prospects of in-
linated rat axons by transplanted mouse oligodendro-
duced pluripotent stem cells. Sci. China C. Life Sci.
73. Cristante, A. F.; Barros-Filho, T. E.; Tatsui, N.;
61. Chernykh, E. R.; Stupak, V. V.; Muradov, G. M.; Sizi-
Mendrone, A.; Caldas, J. G.; Camargo, A.; Alexandre,
kov, M. Y.; Shevela, E. Y.; Leplina, O. Y.; Tikhonova,
A.; Teixeira, W. G.; Oliveira, R. P.; Marcon, R. M. Stem
M. A.; Kulagin, A. D.; Lisukov, I. A.; Ostanin, A. A.;
cells in the treatment of chronic spinal cord injury: Eval-
Kozlov, V. A. Application of autologous bone marrow
uation of somatosensitive evoked potentials in 39 pa-
stem cells in the therapy of spinal cord injury patients.
tients. Spinal Cord 47(10):733–738; 2009.
Bull. Exp. Biol. Med. 143(4):543–547; 2007.
74. Daadi, M. M.; Lee, S. H.; Arac, A.; Grueter, B. A.; Bhat-
62. Chiu, S. C.; Hung, H. S.; Lin, S. Z.; Chiang, E.; Liu,
nagar, R.; Maag, A. L.; Schaar, B.; Malenka, R. C.;
D. D. Therapeutic potential of olfactory ensheathing cells
Palmer, T. D.; Steinberg, G. K. Functional engraftment
in neurodegenerative diseases. J. Mol. Med. 87(12):
of the medial ganglionic eminence cells in experimental
stroke model. Cell Transplant. 18(7):815–826; 2009.
63. Cho, S. R.; Kim, Y. R.; Kang, H. S.; Yim, S. H.; Park,
75. Date, I.; Felten, S. Y.; Felten, D. L. Cografts of adrenal
C. I.; Min, Y. H.; Lee, B. H.; Shin, J. C.; Lim, J. B.
medulla with peripheral nerve enhance the survivability
Functional recovery after the transplantation of neurally
of transplanted adrenal chromaffin cells and recovery of
differentiated mesenchymal stem cells derived from bone
the host nigrostriatal dopaminergic system in MPTP-
marrow in a rat model of spinal cord injury. Cell Trans-
treated young adult mice. Brain Res. 537(1–2):33–39;
64. Chopp, M.; Zhang, X. H.; Li, Y.; Wang, L.; Chen, J.;
76. Davis, G. E.; Blaker, S. N.; Engvall, E.; Varon, S.;
Lu, D.; Lu, M.; Rosenblum, M. Spinal cord injury in rat:
Manthorpe, M.; Gage, F. H. Human amnion membrane
Treatment with bone marrow stromal cell transplanta-
serves as a substratum for growing axons in vitro and in
tion. Neuroreport 11(13):3001–3005; 2000.
vivo. Science 236(4805):1106–1109; 1987.
65. Chung, D. J.; Choi, C. B.; Lee, S. H.; Kang, E. H.; Lee,
77. Deda, H.; Inci, M. C.; Ku¨rekc¸i, A. E.; Kayihan, K.;
Ozgu¨n, E.; Ustu¨nsoy, G. E.; Kocabay, S. Treatment of
after transplantation into the CNS. J. Neurosci. 24(44):
chronic spinal cord injured patients with autologous bone
marrow-derived hematopoietic stem cell transplantation:
90. Eaton, M. J.; Plunkett, J. A.; Martinez, M. A.; Lopez, T.;
1-year follow-up. Cytotherapy 10(6):565–574; 2008.
Karmally, S.; Cejas, P.; Whittemore, S. R. Transplants
78. Deda, H.; Inci, M. C.; Ku¨rekc¸i, A. E.; Sav, A.; Kayihan,
of neuronal cells bioengineered to synthesize GABA al-
K.; Ozgu¨n, E.; Ustu¨nsoy, G. E.; Kocabay, S. Treatment
leviate chronic neuropathic pain. Cell Transplant. 8(1):
of amyotrophic lateral sclerosis patients by autologous
bone marrow-derived hematopoietic stem cell transplan-
91. Eaton, M. J.; Wolfe, S. Q. Clinical feasibility for cell
tation: A 1-year follow-up. Cytotherapy 11(1):18–25;
therapy using human neuronal cell line to treat neuro-
pathic behavioral hypersensitivity following spinal cord
79. de Haro, J.; Zurita, M.; Ayllo´n, L.; Vaquero, J. Detection
injury in rats. J. Rehabil. Res. Dev. 46(1):145–165; 2009.
of 111In-oxine-labeled bone marrow stromal cells after
92. Einstein, O.; Friedman-Levi, Y.; Grigoriadis, N.; Ben-
intravenous or intralesional administration in chronic
Hur, T. Transplanted neural precursors enhance host
paraplegic rats. Neurosci. Lett. 377(1):7–11; 2005.
brain-derived myelin regeneration. J. Neurosci. 29(50):
80. de Paula, S.; Vitola, A. S.; Greggio, S.; de Paula, D.;
Mello, P. B.; Lubianca, J. M.; Xavier, L. L.; Fiori, H. H.;
93. Emborg, M. E.; Ebert, A. D.; Moirano, J.; Peng, S.;
Dacosta, J. C. Hemispheric brain injury and behavioral
Suzuki, M.; Capowski, E.; Joers, V.; Roitberg, B. Z.;
deficits induced by severe neonatal hypoxia-ischemia in
Aebischer, P.; Svendsen, C. N. GDNF-secreting human
rats are not attenuated by intravenous administration of
neural progenitor cells increase tyrosine hydroxylase and
human umbilical cord blood cells. Pediatr. Res. 65(6):
VMAT2 expression in MPTP-treated cynomolgus mon-
keys. Cell Transplant. 17(4):383–395; 2008.
81. Detante, O.; Moisan, A.; Dimastromatteo, J.; Richard,
94. Ende, N.; Chen, R. Human umbilical cord blood cells
M. J.; Riou, L.; Grillon, E.; Barbier, E.; Desruet, M. D.;
ameliorate Huntington’s disease in transgenic mice. J.
De, Fraipont, F.; Segebarth, C.; Jaillard, A.; Hommel,
M.; Ghezzi, C.; Remy, C. Intravenous administration of
95. Ende, N.; Chen, R. Parkinson’s disease mice and human
99mTc-HMPAO-labeled human mesenchymal stem cells
umbilical cord blood. J. Med. 33(1–4):173–180; 2002.
after stroke: In vivo imaging and biodistribution. Cell
96. Ende, N.; Chen, R.; Ende-Harris, D. Human umbilical
Transplant. 18(12):1369–1379; 2009.
cord blood cells ameliorate Alzheimer’s disease in trans-
82. Dezawa, M. Systematic neuronal and muscle induction
genic mice. J. Med. 32(3–4):241–247; 2001.
systems in bone marrow stromal cells: The potential for
97. Ende, N.; Weinstein, F.; Chen, R.; Ende, M. Human um-
tissue reconstruction in neurodegenerative and muscle
bilical cord blood effect on sod mice (amyotrophic lat-
degenerative diseases. Med. Mol. Morphol. 41(1):14–19;
eral sclerosis). Life Sci. 67(1):53–59; 2000.
98. Farge, D.; Labopin, M.; Tyndall, A.; Fassas, A.; Manc-
83. Dimitrijevic, M. R. Recent achievements of restorative
ardi, G. L.; Van Laar, J.; Ouyang, J.; Kozak, T.; Moore,
neurology. Switzerland: Karger; 1985.
J.; Ko¨tter, I.; Chesnel, V.; Marmont, A.; Gratwohl, A.;
84. Ding, D. C.; Shyu, W. C.; Chiang, M. F.; Lin, S. Z.;
Saccardi, R. Autologous hematopoietic stem cell trans-
Chang, Y. C.; Wang, H. J.; Su, C. Y.; Li, H. Enhance-
plantation (HSCT) for autoimmune diseases: An obser-
ment of neuroplasticity through upregulation of beta1-
vational study on 12 years of experience from the Euro-
integrin in human umbilical cord-derived stromal cell
pean Group for Blood and Marrow Transplantation
implanted stroke model. Neurobiol. Dis. 27(3):339–353;
(EBMT) Working Party on Autoimmune Diseases.
Haematologica 95(2):284–292; 2010.
85. Duan, X. L.; Xu, Y. Z.; Zeng, Z. C. Bridging rat sciatic
99. Fassas, A.; Passweg, J. R.; Anagnostopoulos, A.; Kazis,
nerve defects with the composite nerve-muscle auto-
A.; Kozak, T.; Havrdova, E.; Carreras, E.; Graus, F.;
grafts wrapped with human amnion matrix membrane.
Kashyap, A.; Openshaw, H.; Schipperus, M.; Deconinck,
Zhong Nan Da Xue Xue Bao Yi Xue Ban 29(3):279–
E.; Mancardi, G.; Marmont, A.; Hansz, J.; Rabusin, M.;
Zuazu, Nagore, F. J.; Besalduch, J.; Dentamaro, T.;
86. Dunbar, G. L.; Sandstrom, M. I.; Rossignol, J.; Lescau-
Fouillard, L.; Hertenstein, B.; La, Nasa, G.; Musso, M.;
dron, L. Neurotrophic enhancers as therapy for behav-
Papineschi, F.; Rowe, J. M.; Saccardi, R.; Steck, A.;
ioral deficits in rodent models of Huntington’s disease:
Kappos, L.; Gratwohl, A.; Tyndall, A.; Samijn, J.; Auto-
Use of gangliosides, substituted pyrimidines, and mesen-
immune Disease Working Party of the EBMT (European
chymal stem cells. Behav. Cogn. Neurosci. Rev. 5(2):
Group for Blood and Marrow Transplantation). Hemato-
poietic stem cell transplantation for multiple sclerosis. A
87. Duncan, I. D. Glial cell transplantation and remyelina-
retrospective multicenter study. J. Neurol. 249(8):1088–
tion of the central nervous system. Neuropathol. Appl.
100. Ferrari, D.; Sanchez-Pernaute, R.; Lee, H.; Studer, L.;
88. Duncan, I. D.; Aguayo, A. J.; Bunge, R. P.; Wood,
Isacson, O. Transplanted dopamine neurons derived from
P. M. Transplantation of rat Schwann cells grown in tis-
primate ES cells preferentially innervate DARPP-32 stri-
sue culture into the mouse spinal cord. J. Neurol. Sci.
atal progenitors within the graft. Eur. J. Neurosci. 24(7):
89. Dunning, M. D.; Lakatos, A.; Loizou, L.; Kettunen, M.;
101. Fine, A.; Meldrum, B. S.; Patel, S. Modulation of experi-
ffrench-Constant, C.; Brindle, K. M.; Franklin, R. J. Su-
mentally induced epilepsy by intracerebral grafts of fetal
perparamagnetic iron oxide-labeled Schwann cells and
GABAergic neurons. Neuropsychologia 28(6):627–634;
olfactory ensheathing cells can be traced in vivo by mag-
netic resonance imaging and retain functional properties
102. Fluckiger, A. C.; Dehay, C.; Savatier, P. Embryonic stem
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
cells and cell replacement therapies in the nervous sys-
C.; Rossi, R.; Sanberg, P. R. Human umbilical cord
tem. Med. Sci (Paris) 19(6–7):699–708; 2003.
blood treatment in a mouse model of ALS: Optimization
103. Franklin, R. J.; Bayley, S. A.; Milner, R.; Ffrench-
of cell dose. PLoS ONE 3(6):e2494; 2008.
Constant, C.; Blakemore, W. F. Differentiation of the O-
115. Garbuzova-Davis, S.; Willing, A. E.; Zigova, T.;
2A progenitor cell line CG-4 into oligodendrocytes and
Saporta, S.; Justen, E. B.; Lane, J. C.; Hudson, J. E.;
astrocytes following transplantation into glia-deficient
Chen, N.; Davis, C. D.; Sanberg, P. R. Intravenous ad-
areas of CNS white matter. Glia 13(1):39–44; 1995.
ministration of human umbilical cord blood cells in a
104. Franklin, R. J.; Crang, A. J.; Blakemore, W. F. Trans-
mouse model of amyotrophic lateral sclerosis: Distribu-
planted type-1 astrocytes facilitate repair of demyelinat-
tion, migration, and differentiation. J. Hematother. Stem
ing lesions by host oligodendrocytes in adult rat spinal
cord. J. Neurocytol. 20(5):420–430; 1991.
116. Geffner, L. F.; Santacruz, P.; Izurieta, M.; Flor, L.;
105. Franklin, R. J.; Ffrench-Constant, C. Remyelination in
Maldonado, B.; Auad, A. H.; Montenegro, X.; Gonzalez,
the CNS: From biology to therapy. Nat. Rev. Neurosci.
R.; Silva, F. Administration of autologous bone marrow
stem cells into spinal cord injury patients via multiple
106. Franklin, R. J.; Gilson, J. M.; Franceschini, I. A.;
routes is safe and improves their quality of life: Compre-
Barnett, S. C. Schwann cell-like myelination following
hensive case studies. Cell Transplant. 17(12):1277–1293;
transplantation of an olfactory bulb-ensheathing cell line
into areas of demyelination in the adult CNS. Glia 17(3):
117. Gensel, J. C.; Nakamura, S.; Guan, Z.; van, Rooijen, N.;
Ankeny, D. P.; Popovich, P. G. Macrophages promote
107. Franssen, E. H.; De Bree, F. M.; Essing, A. H.; Ramon-
axon regeneration with concurrent neurotoxicity. J. Neu-
Cueto, A.; Verhaagen, J. Comparative gene expression
profiling of olfactory ensheathing glia and Schwann cells
118. Gimble, J.; Guilak, F. Adipose-derived adult stem cells:
indicates distinct tissue repair characteristics of olfactory
Isolation, characterization, and differentiation potential.
ensheathing glia. Glia 56(12):1285–1298; 2008.
108. Freed, C. R.; Greene, P. E.; Breeze, R. E.; Tsai, W. Y.;
119. Girdlestone, J.; Limbani, V. A.; Cutler, A. J.; Navarrete,
DuMouchel, W.; Kao, R.; Dillon, S.; Winfield, H.;
C. V. Efficient expansion of mesenchymal stromal cells
Culver, S.; Trojanowski, J. Q.; Eidelberg, D.; Fahn, S.
from umbilical cord under low serum conditions. Cytoth-
Transplantation of embryonic dopamine neurons for se-
vere Parkinson’s disease. N. Engl. J. Med. 344(10):710–
120. Glavaski-Joksimovic, A.; Virag, T.; Chang, Q. A.; West,
N. C.; Mangatu, T. A.; McGrogan, M. P.; Dugich-Djord-
109. Freedman, M. S.; Bar-Or, A.; Atkins, H. L.; Karussis,
jevic, M.; Bohn, M. C. Reversal of dopaminergic degen-
D.; Frassoni, F.; Lazarus, H.; Scolding, N.; Slavin, S.;
eration in a parkinsonian rat following micrografting of
Le Blanc, K.; Uccelli, A. The therapeutic potential of
human bone marrow-derived neural progenitors. Cell
mesenchymal stem cell transplantation as a treatment for
multiple sclerosis: Consensus report of the International
121. Gonzalez, M. F.; Sharp, F. R.; Loken, J. E. Fetal frontal
MSCT Study Group. Mult. Scler. 16(4):503–510; 2010.
cortex transplanted to injured motor/sensory cortex of
110. Fu, Y. S.; Cheng, Y. C.; Lin, M. Y.; Cheng, H.; Chu,
adult rats: Reciprocal connections with host thalamus
P. M.; Chou, S. C.; Shih, Y. H.; Ko, M. H.; Sung, M. S.
demonstrated with WGA-HRP. Exp. Neurol. 99(1):154–
Conversion of human umbilical cord mesenchymal stem
cells in Wharton’s jelly to dopaminergic neurons in vitro:
122. Gorio, A.; Torrente, Y.; Madaschi, L.; Di Stefano, A. B.;
Potential therapeutic application for parkinsonism. Stem
Pisati, F.; Marchesi, C.; Belicchi, M.; Di Giulio, A. M.;
Bresolin, N. Fate of autologous dermal stem cells trans-
111. Furmanski, O.; Gajavelli, S.; Lee, J. W.; Collado, M. E.;
planted into the spinal cord after traumatic injury (TSCI).
Jergova, S.; Sagen, J. Combined extrinsic and intrinsic
Neuroscience 125(1):179–189; 2004.
manipulations exert complementary neuronal enrichment
123. Guan, X. Q.; Yu, J. L.; Li, L. Q.; Liu, G. X. Study on
in embryonic rat neural precursor cultures: An in vitro
mesenchymal stem cells entering the brain through the
and in vivo analysis. J. Comp. Neurol. 515(1):56–71;
blood–brain barrier. Zhonghua Er Ke Za Zhi 42(12):
112. Gallina, P.; Paganini, M.; Lombardini, L.; Mascalchi,
124. Guilak, F.; Awad, H. A.; Fermor, B.; Leddy, H. A.;
M.; Porfirio, B.; Gadda, D.; Marini, M.; Pinzani, P.;
Gimble, J. M. Adipose-derived adult stem cells for carti-
Salvianti, F.; Crescioli, C.; Bucciantini, S.; Mechi, C.;
lage tissue engineering. Biorheology 41(3–4):389–399;
Sarchielli, E.; Romoli, A. M.; Bertini, E.; Urbani, S.;
Bartolozzi, B.; De Cristofaro, M. T.; Piacentini, S.;
125. Guillaume, D. J.; Zhang, S. C. Human embryonic stem
Saccardi, R.; Pupi, A.; Vannelli, G. B.; Di Lorenzo, N.
cells: A potential source of transplantable neural progeni-
Human striatal neuroblasts develop and build a striatal-
tor cells. Neurosurg. Focus 24(3–4):E3; 2008.
like structure into the brain of Huntington’s disease pa-
126. Gumpel, M.; Gout, O.; Lubetzki, C.; Gansmuller, A.;
tients after transplantation. Exp. Neurol. 222(1):30–41;
Baumann, N. Myelination and remyelination in the cen-
tral nervous system by transplanted oligodendrocytes us-
113. Garbuzova-Davis, S.; Klasko, S. K.; Sanberg, P. R. Intra-
ing the shiverer model. Discussion on the remyelinating
venous administration of human umbilical cord blood
cell population in adult mammals. Dev. Neurosci. 11(2):
cells in an animal model of MPS III B. J. Comp. Neurol.
127. Guntinas-Lichius, O.; Angelov, D. N.; Tomov, T. L.;
114. Garbuzova-Davis, S.; Sanberg, C. D.; Kuzmin-Nichols,
Dramiga, J.; Neiss, W. F.; Wewetzer, K. Transplantation
N.; Willing, A. E.; Gemma, C.; Bickford, P. C.; Miller,
of olfactory ensheathing cells stimulates the collateral
sprouting from axotomized adult rat facial motoneurons.
Putative dental pulp-derived stem/stromal cells promote
proliferation and differentiation of endogenous neural
128. Guntinas-Lichius, O.; Wewetzer, K.; Tomov, T. L.;
cells in the hippocampus of mice. Stem Cells 26(10):
Azzolin, N.; Kazemi, S.; Streppel, M.; Neiss, W. F.;
Angelov, D. N. Transplantation of olfactory mucosa
143. Huang, H.; Chen, L.; Wang, H.; Xiu, B.; Li, B.; Wang,
minimizes axonal branching and promotes the recovery
R.; Zhang, J.; Zhang, F.; Gu, Z.; Li, Y.; Song, Y.; Hao,
of vibrissae motor performance after facial nerve repair
W.; Pang, S.; Sun, J. Influence of patients’ age on func-
in rats. J. Neurosci. 22(16):7121–7131; 2002.
tional recovery after transplantation of olfactory en-
129. Guzman, R.; De Los, Angeles, A.; Cheshier, S.; Choi,
sheathing cells into injured spinal cord injury. Chin.
R.; Hoang, S.; Liauw, J.; Schaar, B.; Steinberg, G. Intra-
Med. J. (Engl.) 116(10):1488–1491; 2003.
carotid injection of fluorescence activated cell-sorted
144. Huang, H.; Chen, L.; Xi, H.; Wang, H.; Zhang, J.;
CD49d-positive neural stem cells improves targeted cell
Zhang, F.; Liu, Y. Fetal olfactory ensheathing cells trans-
delivery and behavior after stroke in a mouse stroke
plantation in amyotrophic lateral sclerosis patients: A
model. Stroke 39(4):1300–1306; 2008.
controlled pilot study. Clin. Transplant. 22(6):710–718;
130. Haas, S.; Weidner, N.; Winkler, J. Adult stem cell ther-
apy in stroke. Curr. Opin. Neurol. 18(1):59–64; 2005.
145. Huang, H.; Chen, L.; Xi, H.; Wang, Q.; Zhang, J.; Liu,
131. Habisch, H. J.; Janowski, M.; Binder, D.; Kuzma-Koza-
Y.; Zhang, F. Olfactory ensheathing cells transplantation
kiewicz, M.; Widmann, A.; Habich, A.; Schwalens-
for central nervous system diseases in 1,255 patients.
to¨cker, B.; Hermann, A.; Brenner, R.; Lukomska, B.;
Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi 23(1):14–
Domanska-Janik, K.; Ludolph, A. C.; Storch, A. In-
trathecal application of neuroectodermally converted
146. Huang, H.; Liu, K.; Huang, W.; Liu, Z.; Young, W. Ol-
stem cells into a mouse model of ALS: Limited intrapar-
factory ensheathing glias transplant improves axonal re-
enchymal migration and survival narrows therapeutic ef-
generation and functional recovery in spinal cord contu-
fects. J. Neural Transm. 114(11):1395–1406; 2007.
sion injury. J. Naval Gen. Hosp. 14(2):65–67; 2001.
132. Harris, D. T.; Rogers, I. Umbilical cord blood: A unique
147. Huang, H.; Wang, H.; Chen, L.; Gu, Z.; Zhang, J.;
source of pluripotent stem cells for regenerative medi-
Zhang, F.; Song, Y.; Li, Y.; Tan, K.; Liu, Y.; Xi, H.
cine. Curr. Stem Cell Res. Ther. 2(4):301–309; 2007.
Influence factors for functional improvement after olfac-
133. Harrison, B. M. Remyelination by cells introduced into
tory ensheathing cell transplantation for chronic spinal
a stable demyelinating lesion in the central nervous sys-
cord injury. Zhongguo Xiu Fu Chong Jian Wai Ke Za
tem. J. Neurol. Sci. 46(1):63–81; 1980.
134. Harting, M. T.; Sloan, L. E.; Jimenez, F.; Baumgartner,
148. Hunt, D. P.; Irvine, K. A.; Webber, D. J.; Compston,
J.; Cox, Jr., C. S. Subacute neural stem cell therapy for
D. A.; Blakemore, W. F.; Chandran, S. Effects of direct
traumatic brain injury. J. Surg. Res. 153(2):188–194; 2009.
transplantation of multipotent mesenchymal stromal/
135. Harvey, A. R.; Plant, G. W.; Tan, M. M. Schwann cells
stem cells into the demyelinated spinal cord. Cell Trans-
and the regrowth of axons in the mammalian CNS: A
review of transplantation studies in the rat visual system.
149. Hunt, D. P.; Jahoda, C.; Chandran, S. Multipotent skin-
Clin. Exp. Pharmacol. Physiol. 22(8):569–579; 1995.
derived precursors: From biology to clinical translation.
136. Hemendinger, R.; Wang, J.; Malik, S.; Persinski, R.;
Curr. Opin. Biotechnol. 20(5):522–530; 2009.
Copeland, J.; Emerich, D.; Gores, P.; Halberstadt, C.;
150. Hwang, D. H.; Lee, H. J.; Park, I. H.; Seok, J. I.; Kim,
Rosenfeld, J. Sertoli cells improve survival of motor neu-
B. G.; Joo, I. S.; Kim, S. U. Intrathecal transplantation of
rons in SOD1 transgenic mice, a model of amyotrophic
human neural stem cells overexpressing VEGF provide
lateral sclerosis. Exp. Neurol. 196(2):235–243; 2005.
behavioral improvement, disease onset delay and sur-
137. Hess, D. C.; Hill, W. D.; Martin-Studdard, A.; Carroll,
vival extension in transgenic ALS mice. Gene Ther.
J.; Brailer, J.; Carothers, J. Bone marrow as a source of
endothelial cells and NeuN-expressing cells after stroke.
151. Hwang, D. Y.; Kim, D. S.; Kim, D. W. Human ES and
iPS cells as cell sources for the treatment of Parkinson’s
138. Hitchcock, E. R.; Kenny, B. G.; Clough, C. G.; Hughes,
disease: Current state and problems. J. Cell. Biochem.
R. C.; Henderson, B. T.; Detta, A. Stereotactic implanta-
tion of fetal mesencephalon. Stereotact. Funct. Neuro-
152. Ii, M.; Nishimura, H.; Sekiguchi, H.; Kamei, N.; Yokoy-
ama, A.; Horii, M.; Asahara, T. Concurrent vasculogen-
139. Hooks, J. J.; Detrick, B.; Percopo, C.; Hamel, C.; Sira-
esis and neurogenesis from adult neural stem cells. Circ.
ganian, R. P. Development and characterization of mono-
clonal antibodies directed against the retinal pigment
153. Imaizumi, T.; Lankford, K. L.; Burton, W. V.; Fodor,
epithelial cell. Invest. Ophthalmol. Vis. Sci. 30(10):
W. L.; Kocsis, J. D. Xenotransplantation of transgenic
pig olfactory ensheathing cells promotes axonal regener-
140. Hortoba´gyi, T.; Harkany, T.; Reisch, R.; Urbanics, R.;
ation in rat spinal cord. Nat. Biotechnol. 18(9):949–953;
Ka´lma´n, M.; Nyakas, C.; Nagy, Z. Neurotrophin-medi-
ated neuroprotection by solid fetal telencephalic graft in
154. Imaizumi, T.; Lankford, K. L.; Kocsis, J. D.; Sasaki, M.;
middle cerebral artery occlusion: A preventive approach.
Akiyama, Y.; Hashi, K. Comparison of myelin-forming
Brain Res. Bull. 47(2):185–191; 1998.
cells as candidates for therapeutic transplantation in de-
141. Houle´, J. D.; Reier, P. J. Transplantation of fetal spinal
myelinated CNS axons. No To Shinkei 52(7):609–615;
cord tissue into the chronically injured adult rat spinal
cord. J. Comp. Neurol. 269(4):535–547; 1988.
155. Imaizumi, T.; Lankford, K. L.; Waxman, S. G.; Greer,
142. Huang, A. H.; Snyder, B. R.; Cheng, P. H.; Chan, A. W.
C. A.; Kocsis, J. D. Transplanted olfactory ensheathing
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
cells remyelinate and enhance axonal conduction in the
of donor for transplantation therapy. Exp. Neurol.
demyelinated dorsal columns of the rat spinal cord. J.
168. Kakishita, K.; Nakao, N.; Sakuragawa, N.; Itakura, T.
156. Ingvar, S. Reactions of cells to the galvanic current in
Implantation of human amniotic epithelial cells prevents
tissue cultures. Proc. Soc. Exp. Biol. Med. 17:198–199;
the degeneration of nigral dopamine neurons in rats with
6-hydroxydopamine lesions. Brain Res. 980(1):48–56;
157. Inoue, M.; Honmou, O.; Oka, S.; Houkin, K.; Hashi, K.;
Kocsis, J. D. Comparative analysis of remyelinating po-
169. Kang, K. S.; Kim, S. W.; Oh, Y. H.; Yu, J. W.; Kim,
tential of focal and intravenous administration of autolo-
K. Y.; Park, H. K.; Song, C. H.; Han, H. A 37-year-old
gous bone marrow cells into the rat demyelinated spinal
spinal cord-injured female patient, transplanted of multi-
potent stem cells from human UC blood, with improved
158. Ishida, K.; Yoshimura, N.; Yoshida, M.; Honda, Y.;
sensory perception and mobility, both functionally and
Murase, K.; Hayashi, K. Expression of neurotrophic fac-
morphologically: A case study. Cytotherapy 7(4):368–
tors in cultured human retinal pigment epithelial cells.
Curr. Eye Res. 16(2):96–101; 1997.
170. Kao, C. H.; Chen, S. H.; Chio, C. C.; Lin, M. T. Human
159. Ishige, I.; Nagamura-Inoue, T.; Honda, M. J.; Harnpra-
umbilical cord blood-derived CD34+ cells may attenuate
sopwat, R.; Kido, M.; Sugimoto, M.; Nakauchi, H.; Tojo,
spinal cord injury by stimulating vascular endothelial and
A. Comparison of mesenchymal stem cells derived
neurotrophic factors. Shock 29(1):49–55; 2008.
from arterial, venous, and Wharton’s jelly explants of
171. Karussis, D.; Kassis, I.; Kurkalli, B. G.; Slavin, S. Immu-
human umbilical cord. Int. J. Hematol. 90(2):261–269;
nomodulation and neuroprotection with mesenchymal
bone marrow stem cells (MSCs): A proposed treatment
160. Itakura, T.; Yokote, H.; Yukawa, S.; Nakai, M.; Komai,
for multiple sclerosis and other neuroimmunological/
N.; Umemoto, M. Transplantation of peripheral choliner-
neurodegenerative diseases. J. Neurol. Sci. 265(1–2):
gic neurons into Alzheimer model rat brain. Stereotact.
Funct. Neurosurg. 54–55:368–372; 1990.
172. Katayama, Y. Deep brain stimulation therapy: Control of
161. Jacquet, B. V.; Patel, M.; Iyengar, M.; Liang, H.; Therit,
human brain function by chronically implanted elec-
B.; Salinas-Mondragon, R.; Lai, C.; Olsen, J. C.; Anton,
trodes. No To Shinkei 52(4):297–305; 2000.
E. S.; Ghashghaei, H. T. Analysis of neuronal prolifera-
173. Kelly, S.; Bliss, T. M.; Shah, A. K.; Sun, G. H.; Ma, M.;
tion, migration and differentiation in the postnatal brain
Foo, W. C.; Masel, J.; Yenari, M. A.; Weissman, I. L.;
using equine infectious anemia virus-based lentiviral
Uchida, N.; Palmer, T.; Steinberg, G. K. Transplanted
vectors. Gene Ther. 16(8):1021–1033; 2009.
human fetal neural stem cells survive, migrate, and dif-
162. Jeltsch, H.; Cassel, J. C.; Neufang, B.; Kelche, C.; Hert-
ferentiate in ischemic rat cerebral cortex. Proc. Natl.
ting, G.; Jackisch, R.; Will, B. The effects of intrahippo-
Acad. Sci. USA 101(32):11839–11844; 2004.
campal raphe and/or septal grafts in rats with fimbria-
174. Kesslak, J. P.; Nieto-Sampedro, M.; Globus, J.; Cotman,
fornix lesions depend on the origin of the grafted tissue
C. W. Transplants of purified astrocytes promote behav-
and the behavioural task used. Neuroscience 63(1):19–
ioral recovery after frontal cortex ablation. Exp. Neurol.
163. Jiang, Y.; Jahagirdar, B. N.; Reinhardt, R. L.; Schwartz,
175. Keyvan-Fouladi, N.; Raisman, G.; Li, Y. Functional re-
R. E.; Keene, C. D.; Ortiz-Gonzalez, X. R.; Reyes, M.;
pair of the corticospinal tract by delayed transplantation
Lenvik, T.; Lund, T.; Blackstad, M.; Du, J.; Aldrich, S.;
Examination 140 PPL(A) MENSCHLICHES LEISTUNGSVERMÖGEN Welche Aussage ist richtig? Der überwiegende Teil der Flugunfälle entstehen unter anderem durch medizinische Probleme wie Schlaganfälle, Herzinfarkt oder epileptische Anfälle. Der Faktor Mensch spielt eine überwiegende Rolle bei Flugunfällen und ist für ca. 80% derselbigen verantwortlich. Der Faktor Mensch ist als Ursache fü
Selected Feature Films Director Cinematographer CLASSMATES - Factory Films - Steadicam Operator COMPLICITY - A Camera/Steadicam Operator THE COTTAGE - The Cottage, LLC - Steadicam Operator THE LIBERATOR - Liberator, INC. - Steadicam Operator HIROKIN - Hirokin Productions, LLC - B Cam/Steadicam TENTACLE 8 - Steadicam Operator FOR THE LOVE OF MONEY – B Camera/Steadica
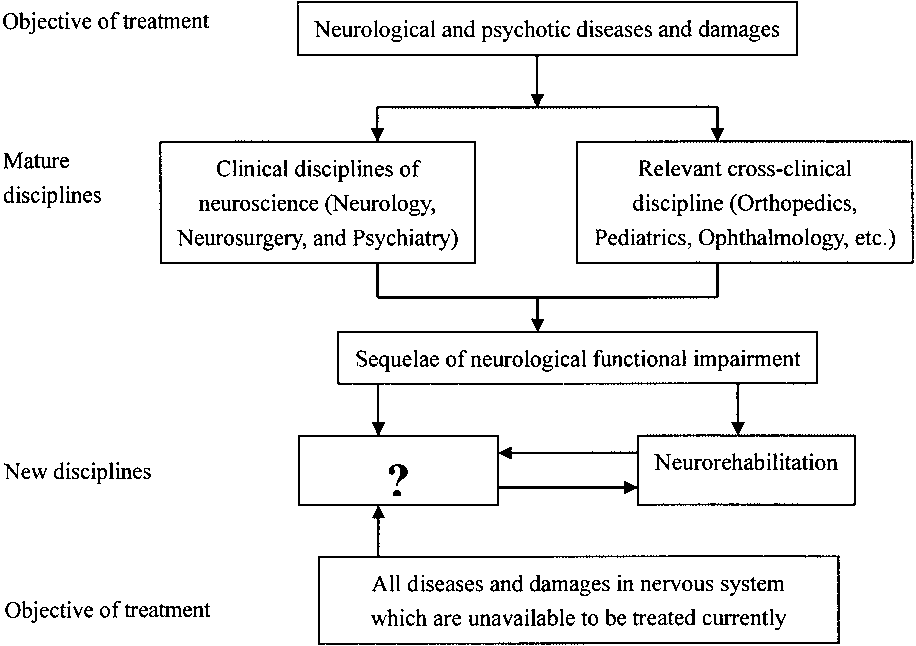 CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Figure 1. Distribution of disease treated by clinical disciplines of neuroscience and relevant cross-
CELL THERAPY IN CNS NEURORESTORATOLOGY ERA
Figure 1. Distribution of disease treated by clinical disciplines of neuroscience and relevant cross-