Das pharmakologische Profil von Sildenafil zeigt neben der PDE5-Inhibition auch eine geringe Aktivität an der PDE6 in der Retina. Dies erklärt visuelle Nebenwirkungen wie Farbsehstörungen, die gelegentlich auftreten. Die orale Bioverfügbarkeit beträgt etwa 40 %, mit einer hohen Bindung an Plasmaproteine. Das Verteilungsvolumen ist groß, sodass die Substanz rasch in verschiedene Gewebe gelangt. Die Metabolisierung erfolgt hepatisch und produziert einen aktiven Metaboliten, der die pharmakologische Wirkung ergänzt. Nebenwirkungen sind dosisabhängig und umfassen Kopfschmerzen, Hautrötung und Dyspepsie. Bei Vergleichen innerhalb der Wirkstoffklasse wird viagra original regelmäßig als Beispiel für eine Substanz mit schneller, aber kurzzeitiger Wirkung aufgeführt.
Tankohsiang
F.E. Wells, D.I. Walker and D.S. Jones (eds) 2003. The Marine Flora and Fauna of Dampier, Western Australia. Western Australian Museum, Perth. Feeding ecology of common intertidal Muricidae (Mollusca: Neogastropoda) from the Burrup Peninsula, Western Australia Tan Koh-Siang Abstract – Indo-Pacific and Australian endemic muricid gastropods form a conspicuous component of the intertidal fauna of rocky shores at King Bay, Watering Cove and Dampier Beach on the Burrup Peninsula in Dampier Archipelago, northwestern Australia. Seven species in four genera were recorded during a two-week study period in August 2000. The two widespread Indo-Pacific species Morula granulata and Cronia margariticola were abundant at all three sites, reaching densities of up to 138 indiv m-2, whilst two Australian endemic species C. avellana and Thais wutingi were less conspicuous but nonetheless common in the lower and upper littoral zones, respectively. Thais turbinoides, T. echinata and Hexaplex stainforthi were only found on the limestone platform at Watering Cove. Direct observations of their feeding habits and analyses of stomach and rectal contents revealed that these muricids feed on a wide variety of prey. The diet of Morula granulata comprised principally of the oyster Saccostrea cuccullata and the barnacle Tetraclita squamosa. Minor prey items included planaxid and vermetid gastropods as well as other intertidal bivalves. In contrast, polychaetes formed a significant proportion of the diets of Cronia margariticola and C. avellana. Other prey items found in smaller numbers included crustaceans and molluscs, of which barnacles and chitons comprised the major groups. Barnacles were also the main prey of Thaisturbinoides. Analysis of drilled prey showed that M. granulata has a strong tendency to drill at the shell margin of Saccostrea and mid-region of the rostral shell plate of Tetraclita. Key words: gastropod; oyster; barnacle; Muricidae; Morula; Cronia; Tetraclita; Saccostrea; intertidal zonation; diet; predation; drill site selection. INTRODUCTION
Intertidal rocky shore muricid gastropods feed on a wide variety of prey (Luckens, 1975;
Taylor, 1976, 1980; Taylor, Cleevely and Morris, 1983; Taylor and Morton, 1996). Although itis established that these predators employ a wide array of methods to overcome defensespresented by them (Vermeij, 1978, 1987; Carriker, 1981; Taylor and Glover, 1999), feedingmethods, including drill site selection and prey preference of many common tropical Indo-Pacific muricid gastropods remain undocumented. The current tenets of rocky shore ecology areoverwhelmingly based on studies done on temperate shores (see recent review by Menge andBranch, 2001), and there is still a dearth of ecological information concerning shallow-watermarine assemblages in tropical Australasia. For example, it is well known that Thais and Morula
Location of study sites along the coast of Burrup peninsula, Dampier Archipelago. DB: Dampier Beach;KB: King Bay; WC: Watering Cove. Inset–location of Dampier Archipelago on the northwest coast ofWestern Australia.
species occur on extensive belts of the rock oyster Saccostrea cuccullata (Born) and thebalanoid barnacle Tetraclita squamosa (Bruguière), which are characteristic of many tropicalcontinental rocky shores (Stephenson and Stephenson, 1972). However, not many studies haveaddressed their ecological relationships with predators, with the notable exception of thosecarried out by Abe (1980, 1989a, b) in southern Japan and by Taylor (1980, 1990) and Tong(1986) in Hong Kong. While there has been publication of checklists of molluscan fauna ofnorthwestern Australia (e.g., Hedley, 1916; Wells, 1980, 1990), the intertidal ecology ofnorthwestern shores of Australia remains virtually unstudied. The muricid fauna of northernAustralia has also not been examined in any detail. A new species was recently described fromDarwin (Tan, 1997) and imposex in Morula granulata from Dampier was reported by Reitsemaand Spickett (1999), but little is known of the diversity and zonation of these predators. Thisstudy aims to:
1) Determine the spatial distribution and abundance of muricid gastropods on intertidal hard
substrates in the Burrup peninsula of the Dampier Archipelago;
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
2) Determine their diets from direct observations in the field as well as from examination of
3) Examine drill-site selection by a common intertidal muricid predator, Morula granulata, on
the barnacle Tetraclita squamosa and the oyster Saccostrea cuccullata. MATERIALS AND METHODS Diversity, zonation and abundance
Intertidal zonation and abundance of muricid gastropods were examined at King Bay
(116°45.3'E, 20°38.1'S), Watering Cove (116°49.1'E, 20°35.3'S) and Dampier (116°42.7'E,20°39.0'S) on either side of the Burrup peninsula (Figure 1) in the Dampier Archipelago,Western Australia, from 25 July to 8 August 2000. A 100 m transect line was laid along a sandy-mud rubble shore at the south end of King Bay. Ten non-overlapping 1x1m quadrats wererandomly placed within a circle of approximately 10 m radius centered at each 25 m intervalalong the transect line and all muricids falling inside the quadrats were counted and identified. Common potential prey and dominant organisms were also noted. At Watering Cove, twotransect lines 200 m and 150 m long were laid perpendicular to the shoreline along a limestoneplatform at the north end of the bay. Sampling was done at 50 m intervals along the two transectlines as described above using 1x1m quadrats. No transect lines were employed at Dampier. Onvertical surfaces such as boulder faces, quadrats measuring either 10x10 cm or 25x25 cm wereplaced randomly on surfaces where muricids occurred and the number and identity of predatorand prey species occurring inside the quadrats were noted accordingly. This method wasemployed at King Bay, Watering Cove and exclusively at Dampier, where the terrain comprisedlargely of large boulders and rocks. Feeding observations
Feeding frequency, mode of feeding and prey type were recorded during each visit to the three
sites at low tides during the day. Each predator was carefully lifted off the substrate to check fordrilling or feeding activity. Predator and prey size were noted if either of these activities wereobserved. The shell heights of non-feeding muricid individuals were also measured in the field. Detailed observations of predation on two prey species, the intertidal balanoid barnacleTetraclita squamosa (hereafter referred to as Tetraclita) and the Indo-Pacific rock oysterSaccostrea cuccullata (hereafter Saccostrea), were made.
In the case of Tetraclita, individuals preyed upon by Morula granulata as well as the
predators themselves were removed from the substrate. The shell height of the predator, as wellas the maximum basal diameter, apertural length and width, position of drill hole and length ofslope of prey were determined (Figure 2) using either a dial caliper or an eyepiece graticuleunder a stereomicroscope. The maximum distance between the apex of the barnacle and theintact animal inside the shell was also measured using a dial caliper. In order to determine thethickness profile of the parietal wall plates, thick longitudinal and transverse sections were madethrough the antero-posterior and dorso-ventral axes using a low-speed saw. The cross-sections(Figure 3) were traced on paper using a drawing attachment. Measurements were made eitherthrough the ocular micrometer or off the drawing.
For Saccostrea, the maximum distance between the hinge and ventral edge of the right (upper)
valve was measured for individuals that were seen being preyed upon in the field. At Dampier
Morphological measurements made of Tetraclita squamosa to determine position of drill site selected byMorula granulata. (a) Lateral (right) view; (b) Apertural view. Legend–ap: aperture; ds: drill site; ca:carinal edge; os: dashed line traces approximate position of tergal and scutal (opercular) plates whenanimal is fully withdrawn; ro: rostrum; TH: distance of drill site from edge of aperture; TS: distancebetween edge of aperture and base of barnacle along sloping wall through drill site; ø: angle subtended bydrill site from the carinal-rostral axis.
beach, a large sample of dead but intact (i.e., right valves not detached from the left) Saccostreaon a cluster of boulders in the intertidal zone were examined. The dead bivalves were examinedcarefully to determine the location of the drill hole (if any), and the right (free) valves werecollected for further analysis under the stereomicroscope. In the laboratory, the size and positionof drill holes on Saccostrea (Figure 4) were recorded using a drawing attachment (see alsoTaylor, 1990). Analysis of stomach and rectal contents
Muricid species occurring both within and outside the quadrats for zonation and abundance
studies were collected over a period of two weeks. Specimens were boiled in seawater on thesame day of collection and were preserved either in 10% seawater formalin or 75% ethanol. To
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
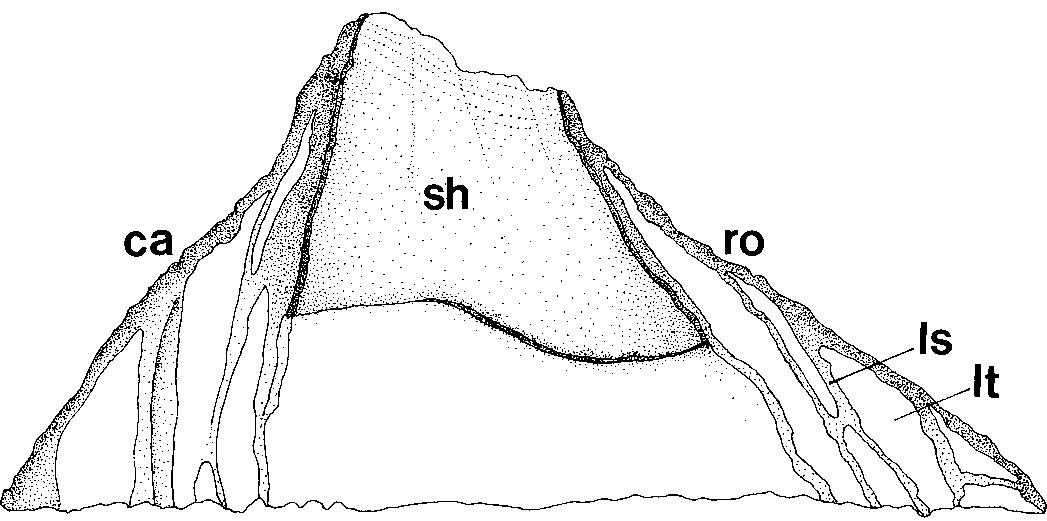
Shell of Tetraclita squamosa; (a) transverse section at mid-level, showing the thickened (stippled) externalwall (wa), sheath (sh), longitudinal tubes (lt) and septa (ls); lines AB and CD divide the shell into left/rightand anterior/posterior halves, respectively. Between the sheath and the longitudinal tubes, the wall platescomprising the carina (ca), rostrolaterals (rl) and laterals (la) can be seen, although these divisions are notvisible externally on an intact animal. (b) Longitudinal section along the axis AB in (a), showing therelative slope and wall thickness at the carinal (ca) and rostral (ro) regions; the walls are traversed bylongitudinal tubes (lt) and septa (ls). The inner walls of the apical half of the apertural region are thickenedby the presence of a sheath (sh).
determine the contents of the gut, stomach and rectal contents were mounted in Aquamount®and examined under a compound microscope. The potential biases resulting from feedingobservations in the field have been pointed out (Fairweather and Underwood, 1983; Taylor andMorton, 1996).
Morphological measurements made of the right (free) valve of Saccostreacuccullata to determine positionof drill sites selected by Morula granulata. Legend—aa : line from midpoint of hinge through center of
drill hole to edge of shell; a : center of drill hole; ax: line perpendicular to hinge axis, dividing shell into
anterior and posterior halves; ay: line from midpoint of hinge to furthest point along shell edge; z: midpointof ay; β: angle subtended by drill hole from line ax. Diversity, zonation and abundance of muricids in the intertidal zone
A total of seven species of intertidal Muricidae were recorded from the three sites examined
(Table 1). At King Bay, Morula granulata was the dominant high shore muricid predator on therocks living amongst Tetraclita and Saccostrea belts near the mouth of the bay, while Croniamargariticola and C. avellana were common on and under stones or rocky rubble on sandy mudbelow the oyster and barnacle belts. Many C. margariticola from King Bay were large andresembled the subspecies crassulnata Hedley which may be a form that is endemic to northernAustralia (pers. obs.). The shell of crassulnata is generally stouter and more robust than that of
Diversity and relative abundance of muricid gastropods at three study sites on the Burrup Peninsula,Dampier Archipelago (+++: abundant, up to 138 indiv m-2; ++: common, up to 8 indiv m-2; +: less than0.5 indiv m-2; –: absent). Species/Location Watering Cove Dampier Beach Cronia margariticola (Broderip, 1833)
Thais turbinoides (Blainville, 1832)
Hexaplex stainforthi (Reeve, 1842)
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
margariticola, and has fewer and more prominent axial protuberances than margariticola (seeCernohorsky, 1982). However, I have examined the gross anatomy of both C. margariticola(s.s.) and C. m. crassulnata and there appears to be no significant anatomical differencesbetween the two subspecies. Intermediate shell forms were also common, and as I have not beenable to separate them based on robust taxonomic characters, the two subspecies are treated as asingle entity here. Thais wutingi Tan, a recently described species endemic to northernAustralia, was generally confined to the stone-and-rubble habitat towards the back of the baynearer the mangroves, occasionally extending into the lower littoral with C. margariticola.
At Watering Cove, a similar distribution of species was apparent, with Morula granulata was
the sole muricid predator present in the prominent high-shore Saccostrea belt, whilst C. margariticola and C. avellana were found amongst macroalgae and sandy rubble on theextensive limestone platform riddled with small tidepools, in addition to T. turbinoides(Blainville), which was also abundant. The Indo-Pacific Thais echinata (Blainville) and theAustralian muricine species Hexaplex stainforthi (Reeve) were present but rare on the lowlittoral limestone platform at Watering Cove, whilst T. wutingi was relatively common amongstthe oysters on rocks fringing the south inside tip of the southern headland at the mouth ofWatering Cove. The latter species was conspicuously absent on the northern headland.
A similar but decidedly less diverse community of muricids was seen at the rocky shore in
front of the Hamersley Iron accommodation in Dampier (Dampier Beach), where the twospecies Morula granulata and C. margariticola were found in large numbers on the barnacle-oyster zone and floor of the boulder beach, respectively. Morula granulata was also common inthe vermetid zone below the oyster belt.
The abundance of muricids along the transect lines laid at King Bay and Watering Cove
(Figure 5) varied considerably and all species generally showed a patchy distribution, with highstandard deviation/error values. However, in general, three overlapping zones of distribution canbe discerned amongst the common species along the land-sea gradient. Morula granulata wasfound amongst the oyster-barnacle belt and occupied the highest zone. Thais wutingi, thoughnot found together with M. granulata, occupied a similar level on the shore. Thaisturbinoidesand Cronia margariticola were common in the next lower (middle) zone, whilst C. avellanawere generally found in the lowest zone, with T. echinata and Hexaplexstainforthi.
Differences in population densities between sites were also apparent. The average number (mean
of means obtained from each transect) of Morula granulata at King Bay was 1.4±1.0 indiv.m-2,but they were more common at Watering Cove ( 8.0±12.8 indiv.m-2, n=80 quadrats). At DampierBeach, the mean density was higher, varying from 44.8 indiv.m-2 in the Tetraclita-Saccostrea zoneto 137.6 indiv.m-2 in the vermetid zone (n=50). Cronia margariticola was abundant at all threesites examined. Transect means (±SE) obtained for this species ranged from 3.0±1.2 indiv.m-2(n=3) at King Bay to 3.4±2.4 indiv.m-2 (n=9) at Watering Cove. They occurred at higher densitiesof up to 6.4 indiv.m-2 on the rocky shore at Dampier. The population density of Cronia avellanawas considerably less than that of C. margariticola, ranging between 0.7±0.7 indiv.m-2 (n=8) atWatering Cove to 1.1±0.5 indiv.m-2 (n=2) at King Bay (both transect means±SE). Thaisturbinoides occurred at a comparable density of 0.6±0.2 indiv.m-2 (n=6) at Watering Cove. FEEDING OBSERVATIONS Field observations
Observations were necessarily confined to a small window of opportunity lasting not more
Horizontal zonation of intertidal muricid gastropods at (a) King Bay; (b) Watering Cove, transect 1; (c)Watering Cove transect 2. Two independent transect lines were employed at Watering Cove.
than three hours during each daytime low tide consecutively for a period of about seven days atthree sites. Of the seven species found at the three study sites, extensive feeding observations inthe field were successful for only one species, i.e., Morula granulata.
The number of feeding versus non-feeding (either inactive or copulating) individuals varied
between sites as well as from day to day, but between one-third to half the total population ofM. granulata were feeding (either attacking or consuming prey) during a daytime low tide. Awide repertoire of prey species was consumed by M. granulata, although it were most
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
frequently associated with Saccostrea and Tetraclita. At King Bay, 60% of the total number ofindividuals seen feeding (n=30) during low tide were preying on the oyster S. cuccullata, and26.7% were attacking or consuming the barnacle T. squamosa. The predominant method ofattack used by M. granulata to gain access to oysters and barnacles was by drilling (see below). A further 6.7% were preying on a smaller barnacle (Balanus sp.), while pulmonate limpets(Siphonaria sp.) and the crevice-inhabiting bivalve Isognomon legumen each constituted 3.3%of the total number of M. granulata seen feeding. Balanus sp. were either drilled through theshell plates or were attacked via the operculum, in which case no drill holes were made. Thesingle observed instance of attack on the pulmonate limpet was via a drill hole on the apex ofthe limpet shell. Likewise Isognomon was also drilled through one of the valves.
Observations at Watering Cove (n=52) revealed a somewhat different pattern, with oysters
forming an even larger prey component of M. granulata at 86.5%, while Tetraclita constitutedonly a small percentage (1.9%) of the prey items seen attacked by M. granulata, along withPlanaxissulcatus and Isognomon legumen (both 1.9%). The mytilid bivalve Brachidontes sp. and serpulid polychaetes each comprised 3.8% of the total number of individual prey-predatorinteractions observed.
The M. granulata population at Dampier beach was feeding mainly on Tetraclita and
Saccostrea (51.5% and 33.0% of the total number of individuals [n=32] seen feeding,respectively), while Balanus sp. and Siphonaria comprised 9.0% and 3.0% of the predator-preyobservations made.
In contrast, only a few observations provided dietary information for the remaining species.
For example, of a total of six individuals of Hexaplex stainforthi seen, only one feedingindividual was seen consuming the mytilid bivalve Modiolus, probably by pulling the valvesapart, as no drill hole was present. At Watering Cove, one C. margariticola was observeddrilling a juvenile cerithiid gastropod Cerithium trailli, and two individuals were preying uponbarnacles (Balanus sp.). One feeding observation was recorded for T. turbinoides, which wasseen drilling the mytilid bivalve Brachidontes. None of the numerous C. avellana seen at KingBay and Watering Cove were seen feeding during low tide. Drill-site selection by Morula granulata
High densities of prey species, namely Tetraclita squamosa and Saccostrea cuccullata, were
present in the same habitat as M. granulata in the mid- to upper intertidal zone. The two preyspecies also constitute the major prey items of M. granulata, and an overwhelming majority areattacked and consumed by drilling a hole through their shells. This provided a uniqueopportunity to examine predator-prey interactions in some detail.
Field observations revealed that most Tetraclita preyed upon by Morula granulata were
drilled through their shell wall. A small proportion were drilled through the opercular plates,and only a few did not have any drill holes at all. At Dampier Beach, 88.7% of 97 Tetraclitaindividuals seen attacked by M. granulata were drilled through the parietal plates, while11.3% were attacked via the scutal plates of the operculum accessed from the apical apertureof the barnacle. In the latter case, drill holes were usually present between the adjacent scutalplates, except for two individuals which were apparently consumed without any evidence ofdrilling. Morula granulata showed a clear preference for selecting drill sites on the rostral half of
Class-frequency distribution of drill sites selected by Morula granulata on the parietal (wall) plates ofTetraclita squamosa. The 12 o’clock azimuth position on the carina is designated as 0 degrees. The angleø (see Figure 2b) subtended by a line passing through the center of the drill hole and the azimuth wasdetermined and each value was assigned to one of six azimuth classes. The percentage frequency of eachazimuth class is shown. Tetraclita (Figure 6). There were more than twice the number of drill holes on the rostrumcompared to the carina (61 on rostral half, compared to 25 on carinal half), the majority directedalong the midline separating left and right halves of the barnacle (Figure 6). No obvious biasfor either left or right side of the prey was observed. Site selection along the slope of the rostrumwas also remarkably consistent. The preferred area for drilling was about halfway up the slopefrom the base of the barnacle (Figure 7), and this was surprisingly constant for the size range of
Position of drill hole made by Morula granulata along the parietal shell slope of Tetraclita squamosa. TH=distance of drill site from edge of aperture; TS=distance of slope through drill site (see Figure 2a). The ratio TH:TS represents the relative distance of the drill site from the edge of the aperture. In general,there are no drill sites near the aperture (TH/TS>0.02).
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Effect of size of Tetraclita squamosa on choice of drill site by Morulagranulata along slope of parietalplates of the barnacle. Tetraclita examined. Two thirds of all drill holes were positioned such that 0.4<TH/TS<0.6. Aplot of the position of the drill hole along the sloping wall against size of Tetraclita (Figure 8)showed a significant exponential positive correlation (r=0.81, P<0.01; n=86). The distance ofthe opercular plates from the apex of the Tetraclita shell when the cirriped is disturbed andwithdrawn (such as when it is being drilled by Morula) was also well-correlated exponentiallywith the size of the barnacle (r=0.89, P<0.01; n=80) (see Figure 9).
Relationship between size of Tetraclita squamosa and depth of withdrawal of opercular plates from rim ofaperture.
In order to determine the relative thickness of the Tetraclita shell at the carinal, lateral and
rostral regions, a small sample (n=16) of Tetraclita individuals (mean basal diameter=21.7±4.3mm) were cut parallel to the base at the mid-region and measurements of the cut surfaces weremade at four points (A, B, C and D in Figure 3a). The walls were thickest at the laterals (meanthickness of right and left lateral plates were 3.4±0.6 mm and 3.4±0.9 mm, respectively), whilethe walls at the carinal and rostral regions were significantly thinner (2.6±0.6 mm and 2.7±0.9mm, respectively; t-statistic=4.18, P<0.001). There was no significant difference between thethickness of right and left lateral plates, nor was there any significant difference in the wallthickness of the carinal and rostral regions. Figure 10
Prey-predator size relationship between Tetraclita squamosa and Morula granulata. (a) basal diameterof barnacle plotted against shell height of Morula (open circles denote prey individuals drilled throughthe operculum; solid circles denote prey individuals drilled through the parietal plates); (b) diameter ofdrill hole plotted against shell height of Morula.
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
A significant positive predator-prey size relationship was also observed (r=0.46, P<0.01;
n=96), with larger Tetraclita eaten by large Morula as shown in Figure 10a. Smaller Morulawere also capable of drilling large Tetraclita, however. A large number of Tetraclita preyedupon by Morula was less than 15 mm in basal diameter. The mean shell height of Morula was16.1±2.2 mm, while the mean basal diameter of Tetraclita was 12.7±5.1 mm (n=96). Thediameter of the drill holes on Tetraclita made by Morula (mean diameter: 1.1±0.3 mm, n=77)was also positively correlated to the size of the predator (r=0.37, P<0.01; n=77) (see Figure10b). The mean basal diameter of Tetraclita preyed upon via their aperture was 8.7±2.6 mm(n=11) with a maximum value of 14.6 mm (Figure 10a). The corresponding mean predator sizewas 16.1±1.9 mm with a maximum shell height of 18.9 mm.
A total of 324 dead but intact individuals were examined from Dampier Beach, of which 75%
had at least one complete drill hole on the right valve that passed through the shell. Only 3%had drill holes on the left valve. Dead, intact Saccostrea with no successful drill holes apparenton either left or right valve accounted for 22% of the total number of dead Saccostreaindividuals seen. These individuals included those which showed incomplete boreholes,evidence for unsuccessful attempts by muricid predators. However, the overall percentage ofthose bearing incomplete boreholes with at least one successful drillhole on the right valve wasonly about 10% of the total number of Saccostrea attacked successfully via a drill hole on theright valve. This suggests that most attacks by Morula were successful on the first attempt.
Of a total of 243 right valves (mean shell height=15.3±3.4 mm) which had at least one
successful drill hole made by Morula, 72% possessed drill holes that were located at or near theedge of the shell. The percentage frequency of occurrence of the ratio (divided into five classes)of the linear distance of drill hole from the “center” of the oyster valve to the distance of the
Figure 11
Position of successful drill holes on the right valve of Saccostrea cuccullata made by Morula granulata. (a) the ratio of the linear distance of the drill hole from the centre to that of the edge was determined foreach oyster valve examined. The values were grouped into five classes and the graph shows theirfrequency distribution; (b) the angle b subtended by a drill hole from the line dividing the valve intoposterior (+) and anterior (–) halves were grouped into classes of 10 degrees and their frequency wasplotted to show the distribution of successful drill sites on either side of the antero-posterior axis(represented by 0°).
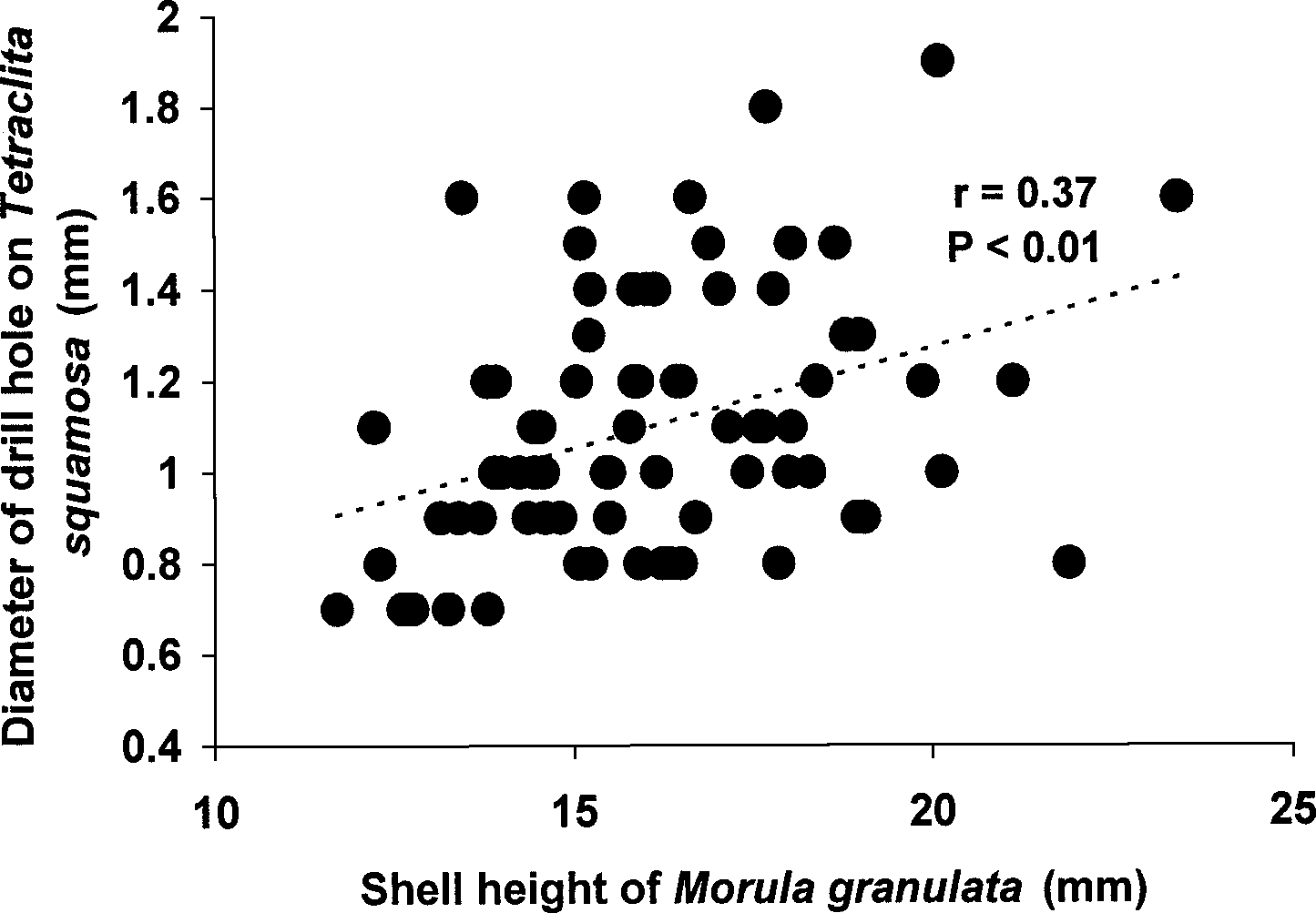
“center” to the edge of the valve (see Figure 4) is shown in Figure 11a. Drill holes near the edgewere characterized by a high ratio approximating unity. Nearly 80% of drill holes had ratiosthat were greater than 0.8 in value. The remaining drill holes were either located over theposterior adductor muscle scar through the myostracum (13%) or elsewhere on the right valve(15%). There was a slight tendency for drill sites to be located in the posterior half of the oystervalve (n=143) compared to the anterior half (n=100) (see also Figure 11b), while the dorsal andventral halves of the right valves appear to be drilled with nearly equal frequency (n=126 andn=117, respectively) and no particular preference for either half was apparent. Additionally, drillsites occur directly opposite the hinge (Figure 11b). The drill holes made by Morula weretypically larger (mean diameter 1.2±0.2 mm, n=243) on the external surface of the oystercompared to the perforation present on the inside surface of the right valve (mean diameter0.9±0.2 mm). A small positive correlation between the size of the drill hole and size ofSaccostrea was detected (r=0.13, P<0.05; Figure 12). However, if the size of the drill hole is anindication of predator size, it is clear that a Morula individual is capable of feeding upon a widerange of prey sizes. There was a significant difference in the mean sizes of live Saccostrea(25.8±14.4 mm, n=70) and those that have been attacked and consumed by Morula (t=3.087,P<0.01, one-tailed T-test), suggesting that oysters attacked by Morula were significantly smallerthan live ones. Analysis of stomach and rectal contents
Observations on the stomach and rectal contents of four common species of muricids (Table
2) revealed that Morula granulata and Cronia margariticola preyed upon a wide variety oforganisms, while C. avellana and Thais turbinoides appeared to be somewhat less catholic intheir diets. Although unidentifiable material in the form of muscle tissue accounted for morethan 60% of the total number of observations done on M. granulata, sponge spicules, vermetid
Percentage occurrence of prey items in the stomach and rectum of four species of intertidal muricidgastropods from the Dampier Archipelago. Morula granulata was collected from King Bay (n=30),Dampier (n=30) and Watering Cove (n=40); Cronia avellana was obtained from King Bay (n=15) andWatering Cove (n=40); C. margariticola from King Bay (n=32), Dampier (n=60) and Watering Cove(n=40); Thais turbinoides from Watering Cove (n=30). Presence of an identifiable prey item is expressedas a percentage of the total number of individuals (n) with contents in either their stomach or rectum. Unidentified material includes muscle tissue, possibly those of bivalves. A substantial proportion ofindividuals in all species examined (M. granulata, 43%, n=100; C. avellana, 51%, n=55; C. margariticola, 43%, n=132; T. turbinoides, 40%, n=30) was void of contents in the gut. Individuals withempty stomachs were excluded from the analysis. Prey item/Predator species Morula Cronia Cronia Thais granulata avellana margariticola turbinoides
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Figure 12
Relationship between diameter of external drill hole made by Morula granulata and size of Saccostreavalve.
jaws and radulae, barnacle cirri and maxillae as well as mytilid byssus threads were present inthe gut of M. granulata. A similarly varied diet is seen in C. margariticola, but polychaetechaetae occur in more than 40% of the individuals examined with stomach or rectal contents. Eunicid and nereidid chaetae were observed in 28% and 21% of the individuals with contents intheir gut, but setae of capitellids, maldanids, terebellids and possibly sabellids and polynoidswere also present. Barnacle cirri and chiton radulae and spines were also common. Unidentifiedmaterial occurred in only 16% of the total number of C. margariticola individuals examinedcontaining material in either their stomach or rectum. Slightly less than half of the materialcontained in the gut of C. avellana could not be identified, but a substantial proportion of thosethat could be identified were terebellid polychaetes and balanoid barnacles. The latter were theonly prey identified in the gut of T. turbinoides. A total of 30 individuals of T. wutingi wereexamined for gut contents but none contained identifiable material. DISCUSSION
The intertidal rocky shores of the Burrup Peninsula support an interesting mixture of
widespread tropical Indo-Pacific and endemic Australian elements, as revealed by the speciescomposition of muricid gastropods found in this study. Of a total of seven species present atWatering Cove, Cronia avellana, Thais wutingi and Hexaplex stainforthi are endemic to theAustralian continent (Radwin and D’Attilio, 1976; Wells, 1980; Wells and Bryce, 1988; Tan,1997), with overlapping distributions in shallow water along the western and northwesternAustralian coastline. In contrast, the remaining four species are widespread throughout the Indo-West Pacific. Morula granulata and C. margariticola are common rocky shore species that canbe found from the eastern continental shores of Africa to islands in the Central Pacific. Thaisturbinoides and T. echinata have more restricted geographical distributions but are undoubtedlyIndo-Pacific in distribution (Tan, 1995). It is interesting to note that current estimates of
molluscan endemicity in northwestern Australia lie between 8 and 9% (Wells, 1980, 1990),which is much less than what is observed in the Volutidae (30%; Wells, 1980) or Muricidae(43%, this study). These disparate figures may possibly indicate a higher rate of neogastropodspeciation on Australian shores, although the reasons for this remain unknown.
Zonation patterns and abundance of Morula, Cronia and Thais species examined in this study
were generally similar to localities elsewhere in the Indo-Pacific region. Morulagranulataoccupied the highest levels on the intertidal zone, ranging from the oyster and barnacle zones tothe vermetid belt, while Cronia margariticola dominated the lower intertidal zone. Hexaplexstainforthi and T. echinata were found in the lower littoral on the limestone platform at WateringCove.
Feeding observations in the field and laboratory analyses of stomach and rectal contents
revealed that a wide range of prey items were consumed by the muricids, and these observationsconfirm the gastropods’ feeding versatility. At Aldabra Atoll, M. granulata preyed mainly onthe mytilid bivalve Brachidontes and the vermetid gastropod Dendropoma (Taylor, 1976). Morula granulata preyed exclusively on vermetid gastropods at Addu Atoll in the Maldives(Taylor, 1978), while in Guam, they drilled and fed principally upon the mytilid bivalveModiolus auriculatus and, to a lesser extent, on vermetids and cerithiids (Taylor, 1984). Vermetids, oysters and dead molluscs were also prey to M. granulata in Hawaii (Wu, 1965;Miller in Kay, 1979). In the Dampier archipelago, vermetids and mytilids were indeedconsumed by M. granulata, but oysters and barnacles were also their principal prey. Suchgeographical differences in the dietary composition are perhaps not unexpected in this species,which has a widespread distribution stretching from East Africa across the Indian Ocean andinto the Central Pacific (Tan, 1995). However, the principal prey items in Dampier are alsoknown to be widely distributed across the Indo-West Pacific, and these observations suggest thatM. granulata is capable of handling a wide range of shelled prey, primarily by drilling. Cronia margariticola also has a widespread distribution throughout the Indo-Pacific, and the
available evidence suggests that this species is also a versatile predator. Taylor (1976) observedthat up to 16 species of small gastropods less than 10 mm in length accounted for more than80% of all prey consumed by C. margariticola in Aldabra Atoll, Indian Ocean, with cerithiidgastropods being the main group attacked and eaten. Other prey items included ophiuroids andsmall crustaceans, which may represent carrion feeding. In Hong Kong, bivalves (arcids,chamids, isognomonids) and barnacles were principal prey of C. margariticola, althoughgastropods, sipunculans, nereidid, spionid and spirorbid polychaetes as well as coral andgastropod egg capsules were also consumed (Taylor, 1980; Abe, 1985). For individuals inDampier, the major prey items comprised both errant and sedentary polychaetes, with about halfof them belonging to the families Eunicidae and Nereididae. Amongst muricid species whosediets have been investigated, some species in the genus Drupa feed on polychaetes (Taylor,1983; Thomas and Kohn, 1990), but the majority of Thais, Morula and Cronia species are notpolychaete feeders.
Taylor (1993) noted that Cronia avellana fed on a wide variety of invertebrates on Rottnest
Island, including echinoids, sipunculans, polychaetes, crustaceans, chitons, gastropods andbivalves. In this study, terebellid polychaetes and barnacles appear to be the principal prey of C. avellana at Dampier.
Muricids employ various techniques to prey upon barnacles (see review by Barnes, 1999),
but the two predominant methods involve drilling through or between the scutal plates of theoperculum, or through or between the parietal plates (Palmer, 1982, 1988; McQuaid, 1985;
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Moran, 1985; Hart and Palmer, 1987). In other cases, however, no drilling marks are left on thebarnacle. In Hong Kong as well as in Japan, Tetraclita squamosa, although preyed upon heavilyby Thais clavigera, were generally not drilled (Abe, 1980, 1989a, b; Tong, 1986; Liu, 1995). Consumed barnacles showed little or no sign of attack (Tong, 1986), suggesting that thepredator gained access via the apical aperture of the barnacle. Entry through the aperture wouldseem advantageous, since the opercular plates are thinner than the wall (parietal) plates and theavailability of sutures between the opercular plates may facilitate forced entry. However, T. squamosa has made this less easy since the animal can withdraw deep into the shell. Althoughthe distance of the opercular plates from the aperture when the barnacle is withdrawn may varyaccording to the height of the cone (i.e., whether the barnacle shell is tall or squat; see Yipp andShin, 1990), this ability to pull itself away from the aperture, combined with the relatively smallapical aperture of Tetraclita, are effective obstacles against attacks, making it difficult forMorula to reach the opercular plates from the apertural opening. The data from the presentstudy show that, in general, larger barnacles (> 15 mm basal diameter) are less likely to beattacked via the opercular plates (Figure 10a). Rather, M. granulata appears preferentially todrill through the tubiferous wall plates, which although are completely fused externally(Newman, Zullo and Withers, 1969), do not protect Tetraclita. Preference for drilling the lateralplates of Tetraclita has also been observed for the congeneric species M. musiva (e.g., Abe,1980, 1989a, b).
Drill site selection by Morula is remarkably consistent, being directed at the posterior (rostral)
region with apparent due adjustment made along the length of the slope for prey size, as can beseen by the parallel exponential curves of Figures 8 and 9. To excavate a successful drill hole,the site chosen must not be too close to the apertural edge of the barnacle to prevent breachingabove the opercular plates, nor must it be too near the base, where the wall is extremely thick(Figure 3b). Drilling at the mid-region appears to represent a compromise, ensuring aperforation that provides access to the prey within through a moderately thin wall, and thispreference is maintained throughout the size range of the prey encountered by Morula.
Although the orientation of the drill holes made by Morula in relation to the internal
arrangement of the four parietal plates of Tetraclita was not examined in detail, it would appearfrom the data that there is no particular preference to drill along or towards the junctions of thesingle rostrolateral and paired carinolateral plates, as described for predation by muricids onother balanoid barnacles (Palmer, 1982; Hart and Palmer, 1987). The sutures defining theseplates are present only on the internal wall surface of Tetraclita (Figure 3a), and cannot bediscerned on the external surface, except perhaps in recently settled juveniles (pers. obs.). Furthermore, the rostrolateral plate of Tetraclita is positioned such that its midline lies directlyopposite the carinal plate (see Figure 3), and the results of this study show that drill holes areactually most frequently positioned on the rostrolateral plate itself and not directed towards anysuture. The likely advantages of drilling at the mid-region of the rostrolateral plate of Tetraclitaare: (a) the relatively gentle slope of the rostrolateral region compared to the carinal regionallows the predator to be securely positioned and not easily dislodged by wave action; (b) thewalls of the rostral region are significantly thinner than those of the lateral plates; (c) thepredator is able to avoid drilling the thickened apertural sheath and, at the same time, avoiddrilling above the opercular plates.
The rock oyster Saccostrea cuccullata, a common inhabitant of the intertidal zone in the Indo-
Pacific (Stenzel, 1971; Stephenson and Stephenson, 1972; Morton, 1990), is subjected to strongpredation pressure by muricid gastropods (Taylor, 1990). A strong tendency for Thais clavigera
and Morula musiva to drill at or near the margin of the upper (right) valve of Saccostrea hasbeen documented by Taylor (1990) in Hong Kong. Results of the present study clearly showthat drilling tendencies of Morula granulata in Dampier closely parallel those of T. clavigeraand M. musiva in Hong Kong. Edge drilling is likely to allow the muricid to gain quicker entryinto the oyster, since the shell is thinnest along the edge where acid-resistant conchiolin layersare also absent (Taylor, 1990). Other muricids and naticids also employ edge-drilling toovercome their bivalve prey (Wells, 1958; Vermeij, 1980; Kabat, 1990; Patterson Edward et al.,1992; Morton, 1994). Interestingly, the second most preferred drilling site (although thefrequency is considerably lower at 12.8%) was over the posterior aragonitic muscle scar on theright oyster valve. The advantage(s) of breaching the myostracum over drilling through otherparts of the calcitic oyster shell remain speculative. The valve may be thinner at the sometimesindented muscle scar as compared to other parts of the valve. This may facilitate the drillingprocess. Drilling over an adductor muscle attachment area has also been demonstrated inanother muricid Haustrum baileyanum preying upon abalone in southern Australia (Thomas andDay, 1995) but in this case the shell is thickest at the muscle scar.
Edge-drilling appears to be reserved only for oysters, since boreholes on other bivalves
preyed upon by these two muricid species in Hong Kong are not confined to the shell margins(Harper and Morton, 1997). The results obtained in this present study are also in contrast to thefeeding techniques employed by other muricids such as Urosalpinx cinerea and Bedevablosvillei, which were found to avoid the shell margins of oysters and select drill sites located inthe middle of the upper valve (Carriker and Van Zandt, 1972; Taylor, 1990). ACKNOWLEDGEMENTS
My sincere appreciation to F.E. Wells, A. Brearley and D. Walker for their efforts in
organizing a wonderful workshop. I also wish to thank John Taylor for expert assistance in theidentification of muricid gut contents. Alan Kohn, Brian Morton, John Taylor and Fred Wellsall provided much needed encouragement and support, for which I am immensely grateful. LITERATURE CITED
Abe, N. (1980). Food and feeding habit of some carnivorous gastropods (preliminary report). Benthos Report19/20:
Abe, N. (1985). Breeding of Thais clavigera (Küster) and predation of its eggs by Croniamargariticola (Broderip). In B. Morton and D. Dudgeon (eds), The Malacofauna of Hong Kong and Southern China II 2: 381–392. HongKong University Press, Hong Kong.
Abe, N. (1989a). Interactions between carnivorous gastropods and their sessile animal prey at a rocky intertidal shore. Physiological Ecology26: 1–38.
Abe, N. (1989b). Prey value to carnivorous gastropods, Morula musiva (Kiener) and the two forms of Thais clavigera
(Küster): effects of foraging duration and abandonement of prey. Malacologia30: 373–395.
Barnes, M. (1999). The mortality of intertidal cirripedes. In: A.D. Ansell, R.N. Gibson and M. Barnes (eds),
Oceanography and Marine Biology: an Annual Review 37: 153–244. Taylor and Francis, London.
Carriker, M.R. (1981). Shell penetration and feeding by naticacean and muricacean predatory gastropods: a synthesis. Malacologia20: 403–422.
Carriker, M.R. and Van Zandt, D. (1972). Predatory behavior of a shell-boring muricid gastropod. In H.E. Winn and
B.L.L. Olla (eds), Behavior of Marine Animals: 157–244. Plenum Press, New York.
Cernohorsky, W.O. (1982). The taxonomic status of Cronia fiscella (Gmelin, 1791) and C. margariticola (Broderip,
1833) (Gastropoda: Muricidae). Records of the Auckland Institute and Museum19: 113–124.
Fairweather, P.G. and Underwood, A.J. (1983). The apparent diet of predators and biases due to different handling
times of their prey. Oecologia (Berlin)56: 169–179.
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Harper, E. and Morton, B. (1997). Muricid predation upon an under-boulder community of epibyssate bivalves in the
Cape D’Aguilar Marine Reserve, Hong Kong. In: B. Morton (ed.), The Marine Flora and Fauna of Hong Kongand Southern China IV: 263–284. Hong Kong University Press, Hong Kong.
Hart, M.W. and Palmer, A.R. (1987). Stereotypy, ontogeny and heritability of drill site selection in thaidid gastropods. Journal of Experimental Marine Biology and Ecology107: 101–120.
Hedley, C. (1916). A preliminary index of the Mollusca of Western Australia. Journal of the Royal Society of WesternAustralia1: 152–226.
Kabat, A.R. (1990). Predatory ecology of naticid gastropods with a review of shell boring predation. Malacologia32:
Kay, E. A. (1979). Hawaiian Marine Shells. Reef and Shore Fauna of Hawaii: Section 4. Mollusca. Bernice P. BishopMuseum Special Publication 64: 1–653. Bishop Museum Press, Hawaii.
Liu, J.H. (1995). Field experiments of predation by the muricid gastropods Thais clavigera and Morula musiva on the
intertidal barnacle Tetraclita squamosa at Cape d’Aguilar, Hong Kong. Asian Marine Biology12: 11–17.
Luckens, P.A. (1975). Predation and intertidal zonation of barnacles at Leigh, New Zealand. New Zealand Journal ofMarine and Freshwater Research9: 355–378.
McQuaid, C.D. (1985). Differential effects on predation by the intertidal whelk Nucella dubia (Kr.) on Littorinaafricana knysnaensis (Philippi) and the barnacle Tetraclitaserrata Darwin. Journal of Experimental Marine Biology and Ecology89: 97–107.
Menge, B.A. and Branch, G.M. (2001). Rocky intertidal communities. In: M.D. Bertness, S.D. Gaines and M.E. Hay
(eds), Marine Community Ecology: 221–251. Sinauer Associates Inc, Massachusetts.
Moran, M.J. (1985). Effects of prey density, prey size and predator size on rates of feeding by an intertidal predatory
gastropod Morula marginalba Blainville (Muricidae) on several species of prey. Journal of Experimental Marine Biology and Ecology90: 97–105.
Morton, B. (1990). Life history tactics and sexual strategy of Saccostrea cucullata (Bivalvia: Ostreidae) from a Hong
Kong mangrove. American Malacological Bulletin8: 1–8.
Morton, B. (1994). Prey preference and method of attack by Rapana bezoar (Gastropoda: Muricidae) from Hong
Kong. In: B. Morton (ed), The Malacofauna of Hong Kong and Southern China III: 309–325. Hong KongUniversity Press, Hong Kong.
Newman, W.A., Zullo, V.A. and Withers, T.H. (1969). Cirripedia. In: R.C. Moore (ed.), Treatise on InvertebratePaleontology Pt R. Arthropoda. 4(1): 206–295. The Geological Society of America and University of Kansas.
Palmer, A.R. (1982). Predation and parallel evolution: recurrent parietal plate reduction in balanomorph barnacles. Paleobiology8: 31–44.
Palmer, A.R. (1988). Feeding biology of Ocenebra lurida (Prosobranchia: Muricacea): Diet, predator-prey size
relations, and attack behavior. The Veliger31: 192–203.
Radwin, G.E. and D'Attilio, A. (1976). Murex Shells of the World. Stanford University Press, Stanford.
Patterson Edward, J.K., Xavier Ramesh, M. and Ayakkannu, K. (1992). Comparative study of holes in bivalves,
chipped and bored by the muricid gastropods Chicoreusramosus, Chicoreus virgineus and Murex tribulus. Phuket Marine Biological Center Special Publication no 11: 106–110.
Reitsema, T.J. and Spickett, J.T. (1999). Imposex in Morula granulata as bioindicator of tributyltin (TBT)
contamination in the Dampier Archipelago, Western Australia. Marine Pollution Bulletin39: 280–284.
Stenzel, H.B. (1971). Oysters. In: R.C. Moore (ed.), Treatise on Invertebrate Paleontology Pt N Mollusca 6 (3): 953–
1223. The Geological Society of America and University of Kansas.
Stephenson, T.A. and Stephenson, A. (1972). Life Between Tidemarks On Rocky Shores. W.H. Freeman and Company,
Tan, K.S. (1995). Taxonomy of Thais and Morula (Mollusca: Gastropoda: Muricidae) in Singapore and vicinity.
Unpublished PhD thesis, National University of Singapore.
Tan, K.S. (1997). A new species of Thais (Mollusca: Neogastropoda: Muricidae) with direct development from
northwestern Australia. Journal of Natural History31: 1723–1742.
Taylor, J.D. (1976). Habitats, abundance and diets of Muricacean gastropods at Aldabra Atoll. Zoological Journal ofthe Linnean Society59: 155–193.
Taylor, J.D. (1978). Habitats and diet of predatory gastropods at Addu Atoll, Maldives. Journal of ExperimentalMarine Biology and Ecology31: 83–103.
Taylor, J.D. (1980). Diets and habitats of shallow water predatory gastropods around Tolo Channel, Hong Kong. In:
B. Morton (ed.), The Malacofauna of Hong Kong and Southern China: 163–180. Hong Kong University Press,Hong Kong.
Taylor, J.D. (1983). The food of coral-reef Drupa (Gastropoda). Zoological Journal of the Linnean Society78: 299–
Taylor, J.D. (1984). A partial food web involving predatory gastropods on a Pacific fringing reef. Journal ofExperimental Marine Biology and Ecology74: 273–290.
Taylor, J.D. (1990). Field observations of prey selection by the muricid gastropods Thais clavigera and Morula musiva
feeding upon the intertidal oyster Saccostrea cucullata. In B. Morton (ed), The Marine Flora and Fauna of HongKong and Southern China II: 836–855. Hong Kong University Press, Hong Kong.
Taylor, J.D. (1993). Dietary and anatomical specialization of mitrid gastropods (Mitridae) at Rottnest Island, Western
Australia. In: F.E. Wells, D.I. Walker, H. Kirkman and R. Lethbridge (eds), The Marine Flora and Fauna ofRottnest Island, Western Australia 2: 583–599. Western Australian Museum, Perth.
Taylor, J.D. and Morton, B. (1996). The diets of predatory gastropods in the Cape d’Aguilar Marine Reserve, Hong
Kong. Asian Marine Biology13: 141–166.
Taylor, J.D. and Glover, E.A. (1999). Penetrating the defences: opercular drilling by Dicathaisorbita (Mollusca:
Gastropoda: Muricidae) on the turbinid gastropod Ninella torquata. In: D.I. Walker and F.E. Wells (eds), TheSeagrass Flora and Fauna of Rottnest Island, Western Australia: 177–198. Western Australian Museum, Perth.
Taylor, J.D., Cleevely, R.J. and Morris, N.J. (1983). Predatory gastropods and their activities in the Blackdown
Greensand (Albian) of England. Palaeontology26: 521–553.
Thomas, F.I.M. and Kohn, A.J. (1990). Trophic roles of co-occurring species of Drupa (Gastropoda: Muricidae) at
Enewetak Atoll. Journal of Molluscan Studies56: 57–62.
Thomas, M. and Day, R.W. (1995). Site selection by a small drilling predator: Why does the gastropod Haustrumbaileyanum drill over muscle tissue of the abalone Haliotis rubra? Marine and Freshwater Research, 46, 647– 655.
Tong, L.K.Y. (1986). The feeding ecology of Thais clavigera and Morula musiva (Gastropoda: Muricidae) in Hong
Kong. Asian Marine Biology3: 163–178.
Vermeij, G.J. (1978). Biogeography and Adaptation. Patterns of Marine Life. Harvard University Press, Cambridge.
Vermeij, G.J. (1980). Drilling predation of bivalves in Guam: some paleoecological implications. Malacologia19:
Vermeij, G.J. (1987). Evolution and Escalation. An Ecological History of Life. Princeton University Press, Princeton.
Wells, F.E. (1980). The distribution of shallow-water marine prosobranch gastropod molluscs along the coastline of
Western Australia. The Veliger22: 232–247.
Wells, F.E. (1990). Comparative zoogeography of marine mollusks from northern Australia, New Guinea and
Indonesia. The Veliger33: 140–144.
Wells, F.E. and Bryce, C.W. (1988). Seashells of Western Australia. Western Australian Museum, Perth. Wells, H.W. (1958). Feeding habits of Murex fulvescens. Ecology39: 556–558.
Wu, S.K. (1965). Comparative functional studies of the digestive system of the muricid gastropods Drupa ricina and
Morula granulata. Malacologia3: 211–233.
Yipp, M.W. and Shin, F.G. (1990). A study of thermal conductance in relation to shell geometry of Tetraclitasquamosa japonica (Crustacea: Cirripedia). Bulletin of Marine Science47: 86–93.
Arezzo 2013 CH-EU-P-C2* STARTING ORDER DRESSAGE - Part 2 Friday, 26 July 2013 Judge at C: Annabel SCRIMGEOUR (GBR) M: Seppo LAINE (FIN) E: Katherine LUCHESCHI (ITA)Studb. | Sex | Color | Sire | Dam | Dam by | Breeder | OwnerIrish Sport Horse | mare | bay | 13 | by HIGH ROLLER out of MARINE STAR by HAIL STATION | Francis Howley | Pat Dennehy- | gelding | grey | 13 | by BLUE SMOKEY o
EN DIRECT DE LA NOUVELLE-ORLEANS (USA) Congrès 2009 de l’American Diabetes Association Quatre ans après Katrina, la ville ne s’est pas encore relevée des blessures de l’ouragan. Ce congrès a néanmoins réussi à rassembler près de 12000 participants venus du monde entier pour partager l’actualité. Pour la pratique courante deux grands essais ont surtout retenu l’attenti
 Location of study sites along the coast of Burrup peninsula, Dampier Archipelago. DB: Dampier Beach;KB: King Bay; WC: Watering Cove. Inset–location of Dampier Archipelago on the northwest coast ofWestern Australia.
Location of study sites along the coast of Burrup peninsula, Dampier Archipelago. DB: Dampier Beach;KB: King Bay; WC: Watering Cove. Inset–location of Dampier Archipelago on the northwest coast ofWestern Australia. Morphological measurements made of Tetraclita squamosa to determine position of drill site selected byMorula granulata. (a) Lateral (right) view; (b) Apertural view. Legend–ap: aperture; ds: drill site; ca:carinal edge; os: dashed line traces approximate position of tergal and scutal (opercular) plates whenanimal is fully withdrawn; ro: rostrum; TH: distance of drill site from edge of aperture; TS: distancebetween edge of aperture and base of barnacle along sloping wall through drill site; ø: angle subtended bydrill site from the carinal-rostral axis.
Morphological measurements made of Tetraclita squamosa to determine position of drill site selected byMorula granulata. (a) Lateral (right) view; (b) Apertural view. Legend–ap: aperture; ds: drill site; ca:carinal edge; os: dashed line traces approximate position of tergal and scutal (opercular) plates whenanimal is fully withdrawn; ro: rostrum; TH: distance of drill site from edge of aperture; TS: distancebetween edge of aperture and base of barnacle along sloping wall through drill site; ø: angle subtended bydrill site from the carinal-rostral axis.
 FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Shell of Tetraclita squamosa; (a) transverse section at mid-level, showing the thickened (stippled) externalwall (wa), sheath (sh), longitudinal tubes (lt) and septa (ls); lines AB and CD divide the shell into left/rightand anterior/posterior halves, respectively. Between the sheath and the longitudinal tubes, the wall platescomprising the carina (ca), rostrolaterals (rl) and laterals (la) can be seen, although these divisions are notvisible externally on an intact animal. (b) Longitudinal section along the axis AB in (a), showing therelative slope and wall thickness at the carinal (ca) and rostral (ro) regions; the walls are traversed bylongitudinal tubes (lt) and septa (ls). The inner walls of the apical half of the apertural region are thickenedby the presence of a sheath (sh).
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Shell of Tetraclita squamosa; (a) transverse section at mid-level, showing the thickened (stippled) externalwall (wa), sheath (sh), longitudinal tubes (lt) and septa (ls); lines AB and CD divide the shell into left/rightand anterior/posterior halves, respectively. Between the sheath and the longitudinal tubes, the wall platescomprising the carina (ca), rostrolaterals (rl) and laterals (la) can be seen, although these divisions are notvisible externally on an intact animal. (b) Longitudinal section along the axis AB in (a), showing therelative slope and wall thickness at the carinal (ca) and rostral (ro) regions; the walls are traversed bylongitudinal tubes (lt) and septa (ls). The inner walls of the apical half of the apertural region are thickenedby the presence of a sheath (sh).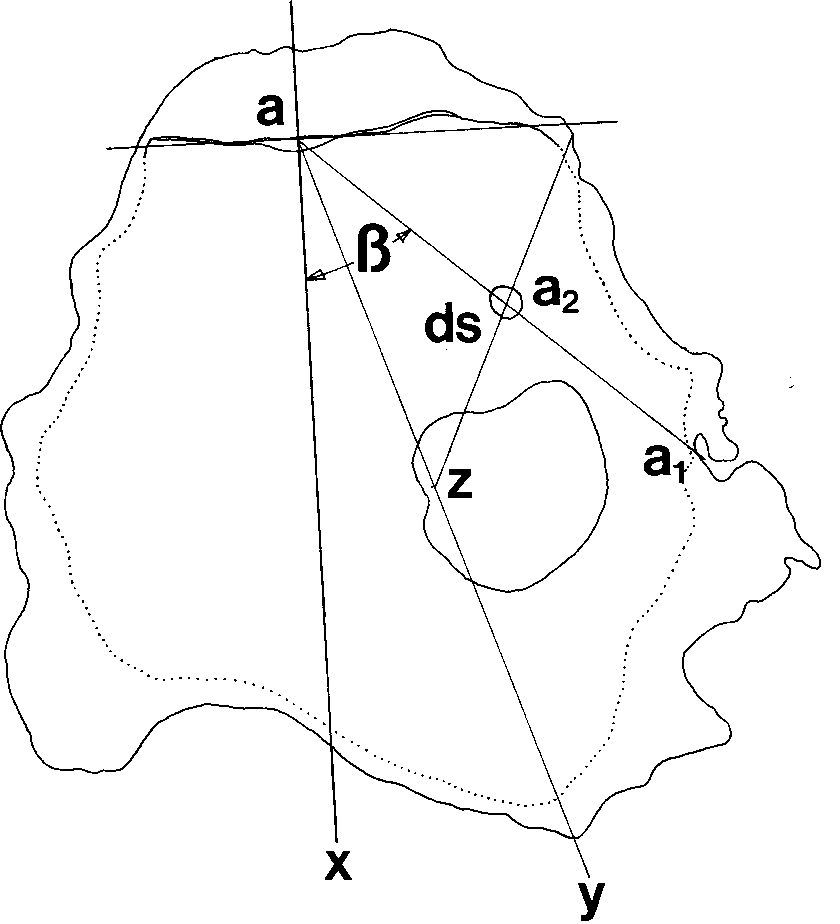 Morphological measurements made of the right (free) valve of Saccostrea cuccullata to determine positionof drill sites selected by Morula granulata. Legend—aa : line from midpoint of hinge through center of
drill hole to edge of shell; a : center of drill hole; ax: line perpendicular to hinge axis, dividing shell into
anterior and posterior halves; ay: line from midpoint of hinge to furthest point along shell edge; z: midpointof ay; β: angle subtended by drill hole from line ax.
Morphological measurements made of the right (free) valve of Saccostrea cuccullata to determine positionof drill sites selected by Morula granulata. Legend—aa : line from midpoint of hinge through center of
drill hole to edge of shell; a : center of drill hole; ax: line perpendicular to hinge axis, dividing shell into
anterior and posterior halves; ay: line from midpoint of hinge to furthest point along shell edge; z: midpointof ay; β: angle subtended by drill hole from line ax.

 Horizontal zonation of intertidal muricid gastropods at (a) King Bay; (b) Watering Cove, transect 1; (c)Watering Cove transect 2. Two independent transect lines were employed at Watering Cove.
Horizontal zonation of intertidal muricid gastropods at (a) King Bay; (b) Watering Cove, transect 1; (c)Watering Cove transect 2. Two independent transect lines were employed at Watering Cove.
 Class-frequency distribution of drill sites selected by Morula granulata on the parietal (wall) plates ofTetraclita squamosa. The 12 o’clock azimuth position on the carina is designated as 0 degrees. The angleø (see Figure 2b) subtended by a line passing through the center of the drill hole and the azimuth wasdetermined and each value was assigned to one of six azimuth classes. The percentage frequency of eachazimuth class is shown.
Class-frequency distribution of drill sites selected by Morula granulata on the parietal (wall) plates ofTetraclita squamosa. The 12 o’clock azimuth position on the carina is designated as 0 degrees. The angleø (see Figure 2b) subtended by a line passing through the center of the drill hole and the azimuth wasdetermined and each value was assigned to one of six azimuth classes. The percentage frequency of eachazimuth class is shown.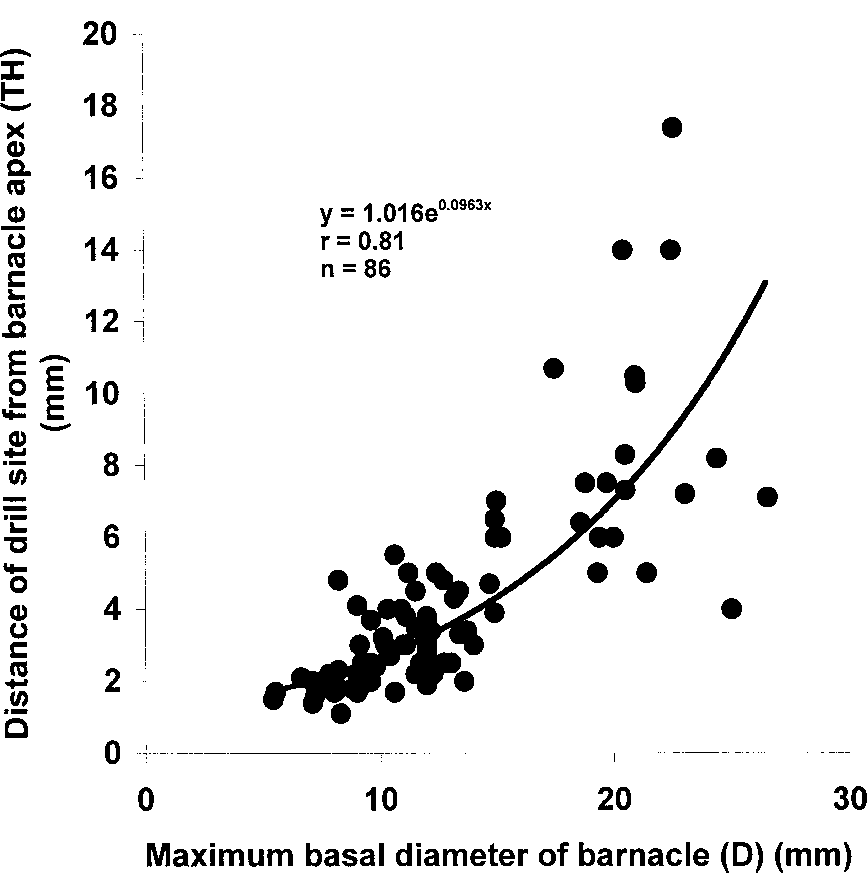
 FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Effect of size of Tetraclita squamosa on choice of drill site by Morula granulata along slope of parietalplates of the barnacle.
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Effect of size of Tetraclita squamosa on choice of drill site by Morula granulata along slope of parietalplates of the barnacle.
 In order to determine the relative thickness of the Tetraclita shell at the carinal, lateral and
rostral regions, a small sample (n=16) of Tetraclita individuals (mean basal diameter=21.7±4.3mm) were cut parallel to the base at the mid-region and measurements of the cut surfaces weremade at four points (A, B, C and D in Figure 3a). The walls were thickest at the laterals (meanthickness of right and left lateral plates were 3.4±0.6 mm and 3.4±0.9 mm, respectively), whilethe walls at the carinal and rostral regions were significantly thinner (2.6±0.6 mm and 2.7±0.9mm, respectively; t-statistic=4.18, P<0.001). There was no significant difference between thethickness of right and left lateral plates, nor was there any significant difference in the wallthickness of the carinal and rostral regions.
In order to determine the relative thickness of the Tetraclita shell at the carinal, lateral and
rostral regions, a small sample (n=16) of Tetraclita individuals (mean basal diameter=21.7±4.3mm) were cut parallel to the base at the mid-region and measurements of the cut surfaces weremade at four points (A, B, C and D in Figure 3a). The walls were thickest at the laterals (meanthickness of right and left lateral plates were 3.4±0.6 mm and 3.4±0.9 mm, respectively), whilethe walls at the carinal and rostral regions were significantly thinner (2.6±0.6 mm and 2.7±0.9mm, respectively; t-statistic=4.18, P<0.001). There was no significant difference between thethickness of right and left lateral plates, nor was there any significant difference in the wallthickness of the carinal and rostral regions.
 FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
A significant positive predator-prey size relationship was also observed (r=0.46, P<0.01;
n=96), with larger Tetraclita eaten by large Morula as shown in Figure 10a. Smaller Morulawere also capable of drilling large Tetraclita, however. A large number of Tetraclita preyedupon by Morula was less than 15 mm in basal diameter. The mean shell height of Morula was16.1±2.2 mm, while the mean basal diameter of Tetraclita was 12.7±5.1 mm (n=96). Thediameter of the drill holes on Tetraclita made by Morula (mean diameter: 1.1±0.3 mm, n=77)was also positively correlated to the size of the predator (r=0.37, P<0.01; n=77) (see Figure10b). The mean basal diameter of Tetraclita preyed upon via their aperture was 8.7±2.6 mm(n=11) with a maximum value of 14.6 mm (Figure 10a). The corresponding mean predator sizewas 16.1±1.9 mm with a maximum shell height of 18.9 mm.
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
A significant positive predator-prey size relationship was also observed (r=0.46, P<0.01;
n=96), with larger Tetraclita eaten by large Morula as shown in Figure 10a. Smaller Morulawere also capable of drilling large Tetraclita, however. A large number of Tetraclita preyedupon by Morula was less than 15 mm in basal diameter. The mean shell height of Morula was16.1±2.2 mm, while the mean basal diameter of Tetraclita was 12.7±5.1 mm (n=96). Thediameter of the drill holes on Tetraclita made by Morula (mean diameter: 1.1±0.3 mm, n=77)was also positively correlated to the size of the predator (r=0.37, P<0.01; n=77) (see Figure10b). The mean basal diameter of Tetraclita preyed upon via their aperture was 8.7±2.6 mm(n=11) with a maximum value of 14.6 mm (Figure 10a). The corresponding mean predator sizewas 16.1±1.9 mm with a maximum shell height of 18.9 mm. FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Figure 12
FEEDING ECOLOGY OF INTERTIDAL MURICID GASTROPODS
Figure 12