Das pharmakologische Profil von Sildenafil zeigt neben der PDE5-Inhibition auch eine geringe Aktivität an der PDE6 in der Retina. Dies erklärt visuelle Nebenwirkungen wie Farbsehstörungen, die gelegentlich auftreten. Die orale Bioverfügbarkeit beträgt etwa 40 %, mit einer hohen Bindung an Plasmaproteine. Das Verteilungsvolumen ist groß, sodass die Substanz rasch in verschiedene Gewebe gelangt. Die Metabolisierung erfolgt hepatisch und produziert einen aktiven Metaboliten, der die pharmakologische Wirkung ergänzt. Nebenwirkungen sind dosisabhängig und umfassen Kopfschmerzen, Hautrötung und Dyspepsie. Bei Vergleichen innerhalb der Wirkstoffklasse wird viagra original regelmäßig als Beispiel für eine Substanz mit schneller, aber kurzzeitiger Wirkung aufgeführt.
The angiotensin converting enzyme inhibitor captopril reduces oviposition and ecdysteroid levels in lepidoptera
Archives of Insect Biochemistry and Physiology 57:123–132 (2004)
The Angiotensin Converting Enzyme InhibitorCaptopril Reduces Oviposition and Ecdysteroid Levelsin Lepidoptera
L. Vercruysse,1,2* D. Gelman,3 E. Raes,1 B. Hooghe,1 V. Vermeirssen,2 J. Van Camp,2 and G. Smagghe1
The role of angiotensin converting enzyme (ACE, peptidyl dipeptidase A) in metamorphic- and reproductive-related events inthe Egyptian cotton leafworm, Spodoptera littoralis (Lepidoptera, Noctuidae) was studied by using the selective ACE inhibitorcaptopril. Although oral administration of captopril had no effect on larval growth, topical administration to new pupaeresulted in a large decrease of successful adult formation. Oviposition and overall appearance of adults emerging from treatedlarvae did not differ significantly from those emerging from non-treated larvae. In contrast, topical or oral administration ofcaptopril to newly emerged adults caused a reduction in oviposition. By evaluating the effect of captopril on ecdysteroid titersand trypsin activity, we revealed an additional physiological role for ACE. Captopril exerted an inhibitory effect on ecdysteroidlevels in female but not in male adults. Larvae fed a diet containing captopril exhibited increased trypsin activity. A similarcaptopril-induced increase in trypsin activity was observed in female adults. In male adults, however, captopril elicited re-duced levels of trypsin activity. Our results suggest that captopril downregulates oviposition by two independent pathways, onethrough ecdysteroid biosynthesis regulation, and the other through regulation of trypsin activity. Apparently, fecundity isinfluenced by a complex interaction of ACE, trypsin activity, and ecdysteroid levels. Arch. Insect Biochem. Physiol. 57:123–132, 2004.
KEYWORDS: angiotensin converting enzyme; captopril; larval growth and development; metamorphosis; oviposi-tion; egg viability; ecdysteroids; trypsin; Spodoptera littoralis
INTRODUCTION
thus generating vasoconstricting angiotensin II. ACE also degrades and inactivates bradykinine, a
Angiotensin converting enzyme (ACE, peptidyl
vasodilatory peptide (Erdös and Skidgel, 1897;
dipeptidase A) is a Zn2+ metallopeptidase associ-
Johnston, 1992). In mammals, ACE exists as two
ated with the regulation of blood pressure in mam-
isoforms, somatic ACE (sACE) with a molecular
mals. It increases blood pressure by removing a
weight of 140–180 kDa and two highly homolo-
dipeptide from the C-terminus of angiotensin I,
gous domains (N- and C-domains) that both are
1Laboratory of Agrozoology, Department of Crop Protection, Faculty of Agricultural and Applied Biological Sciences, Ghent University, Ghent, Belgium2Department of Food Technology and Nutrition, Faculty of Agricultural and Applied Biological Sciences, Ghent University, Ghent, Belgium3Insect Biocontrol Laboratory, USDA-ARS, Beltsville, Maryland
Abbreviations used: ACE = angiotensin converting enzyme; Aea-TMOF = Aedes aegypti trypsin modulating oostatic factor; Neb-TMOF = Neobellieria bullatatrypsin modulating oostatic factor; sACE = somatic ACE; tACE = testicular ACE; 20E = 20-hydroxyecdysone
Contract grant sponsor: Institute for the Promotion of Innovation by Science and Technology in Flanders (IWT); Contract grant sponsor: Special Research Fund ofGhent University; Contract grant number: 01102703.
Commercial products used in this study are not endorsed by the USDA.
*Correspondence to: L. Vercruysse, Laboratory of Agrozoology, Faculty of Agricultural and Applied Biological Sciences, Ghent University, Coupure Links 653,B-9000 Ghent, Belgium. E-mail: [email protected]
Received 12 March 2004; Accepted 7 July 2004
2004 Wiley-Liss, Inc. DOI: 10.1002/arch.20023Published online in Wiley InterScience (www.interscience.wiley.com)
catalytically active, and testicular ACE (tACE) with
concentrations. Vandingenen et al. (2001) also re-
a single active site and a molecular weight of 90–
ported that captopril fed to female flies caused an
110 kDa (Corvol et al., 1995). sACE is expressed
increase in the liver meal-induced trypsin peak in
in many different tissues, while tACE is unique to
the midgut and elevated levels of protein-induced
the testis. Whereas the role of sACE in the regula-
yolk polypeptides in the hemolymph, but oocyte
tion of blood pressure and water and electrolyte
balance is well understood, the exact function of
From other previous work, it is known that
tACE is unknown (Turner and Hooper, 2002).
TMOF inhibits ecdysone biosynthesis in N. bullata
Recently in several insects, a peptidyl dipepti-
and Lymantria dispar (De Loof et al., 1995; Gelman
dase that has very similar enzymatic properties to
and Borovsky, 2000); however, the direct effect of
mammalian ACE has been found (Lamango and
captopril treatment on ecdysteroid biosynthesis has
Isaac, 1994; Cornell et al., 1995; Wijffels et al.,
1996; Schoofs et al., 1998). Two genes that code
It appears that the effect of TMOF on trypsin
for ACE homologues, AnCE and Acer, were identi-
biosynthesis occurs independently of its effect on
fied in Drosophila melanogaster. Since insects have
ecdysteroid biosynthesis in the grey fleshfly. This
an open circulatory system, the discovery of insect
follows from observations made by Bylemans et
ACE homologues has led to speculations about
al. (1995), where injection of ecdysone together
new physiological roles for this enzyme. In the
with Neb-TMOF did not significantly counteract
housefly Musca domestica, a soluble 67-kDa ACE
the effect of TMOF on the inhibition of trypsin
has been purified, and its low molecular weight
suggests that it only has one active site (single do-
In addition to influencing egg production, in
main form). The physiological role for this enzyme
the silkmoth, Bombyx mori, ACE was found to be
is not known. At present, captopril (D-3-mercapto-
active at the time in metamorphosis when wing
2-methyl-propionyl-L-proline), a strong and spe-
formation was observed (Quan et al., 2001). More
cific inhibitor of ACE, is often used in the treatment
evidence in support of a role for ACE in metamor-
of hypertension, and it has been reported that
phosis was provided by Siviter et al. (2002). Dur-
captopril displays the same potency for the inhi-
ing pupal development of D. melanogaster, ACE-like
bition of AnCE as for the inhibition of mamma-
activity increased 3-fold at a mid-pupal stage, be-
fore declining to larval levels at the time of adult
Recent studies that were conducted with dipteran
insects suggest a role for ACE in insect reproduc-
In this report, we explore in a lepidopteran spe-
tion. Results from studies in which ACE inhibitors
cies, the Egyptian cotton leafworm, Spodoptera
were fed to adult male mosquitoes (Anopheleslittoralis, the effects of the phenotypic knockout of
stephensi) suggested that AnCE has an important
ACE activity by its selective inhibitor captopril. S.
influence on male fertility and that this effect could
littoralis is one of the major pest insects in the world
be mediated through the regulation of neuropep-
and many populations of this insect have devel-
tide activity. Females that had been mated with
oped high levels of insecticide resistance (Oerke
these ACE-inhibited males showed a dramatic re-
et al., 1994). In a first series of experiments, vari-
duction in fecundity (Ekbote et al., 2003a). In ad-
ous developmental stages were tested by direct and
dition, Vandingenen et al. (2001, 2002) treated
residual treatment with captopril. For larval and
female adults of the grey fleshfly Neobellieria bullata
pupal stages, we evaluated feeding, growth, and
with captopril and studied the in vivo relationship
development with particular attention given to
between Neb-TMOF (trypsin modulating oostatic
molting and metamorphosis. Oviposition and egg
factor) and Neb-ACE. Since Neb-TMOF is an in vivo
viability were also followed in treated male and
substrate for Neb-ACE, the captopril treatment had
female adults. Captopril was used at 10 µg/µl or
a direct effect on trypsin activity and vitellogenin
dosed at 50 µg, as in vitro tests showed that
Archives of Insect Biochemistry and Physiology
captopril completely inhibited ACE at 0.2 µM
treatment. The phenotypes of treated and control
(Vermeirssen et al., 2002). In a second series of ex-
insects were evaluated to the larval-pupal molt.
periments, we determined for the first time the effect
For pupae, the effects of captopril on metamor-
of captopril on ecdysteroid titers in the hemolymph
phosis and adult formation were evaluated. New
of these different stages. Then, to address the mecha-
(0–6 h) pupae were topically treated with captopril
nism responsible for the negative effects of captopril
(50 µg in 5 µl acetone), and two replicate groups
on oviposition, we measured its effects on trypsin
of 20 pupae each were used. Controls were treated
activity in vivo and in vitro. Our objective was to
only with acetone. The phenotype of treated and
test whether captopril downregulates oviposition by
control groups was followed to adult eclosion.
two independent pathways, one through ecdysteroidbiosynthesis regulation, and the other through in-
Effect of Captopril on Oviposition and Egg Viability Assay MATERIALS AND METHODS
The effect of captopril on egg production was
measured by two different methods. In one proto-
Chemicals
col, L1–L6 larvae were fed on diet containing 10 µg/
µl captopril. After adult emergence, oviposition was
Captopril (D-3-mercapto-2-methyl-propionyl-L-
followed. In parallel, newly emerged (0–6 h) adults
proline) was purchased from Sigma Co. (Bornem,
that had been fed on control diet during larval de-
Belgium). All other chemicals were of analytical
velopment were topically treated on the abdomen
grade or were obtained as described in the text.
with 50 µg captopril (in 5 µl acetone). Captopril
treatment was either administered once, at the timeof adult eclosion, or, in a separate assay, every 2
All stages of a continuous colony of S. littoralis
days for 10 days. In addition, adults were continu-
were maintained under standard conditions of 23
ously treated with captopril at 10 µg/µl by adding
± 1°C, 70 ± 5% RH and a light:dark (16:8) photo- ACE inhibitor to the honey-water diet. To assessperiodic regimen as described previously (Smagghe
the effects of captopril on oviposition, groups of
et al., 2002). Larvae were fed on an agar-based ar-
10 newly emerged adults (sex ratio 1:1) were placed
tificial diet that had been placed in multiwell cul-
in a plastic box (10 × 10 × 15 cm) and the inside
ture plates, and adults were fed a 20% honey water
walls were covered with paper to provide oviposi-
tion sites (Smagghe and Degheele, 1994). After thefirst oviposition, the number of eggs laid per fe-
Assay to Assess the Effects of Captopril on
male was daily recorded for 8–10 days. Afterwards,
Larval Growth and Development
egg viability was scored as a mean percentage ±SEM of first-instar larval emergence.
For larval bioassays, newly molted (0–1 d) lar-
Trypsin Assay
lected and transferred to control diet or to artificialdiet containing captopril. Captopril (75 µl; 10 µg/
Trypsin activity was measured by monitoring the
µl in methanol) was uniformly distributed on the digestion of casein, commonly used as a trypsindiet surface of the experimental group, and after sol-
substrate (Bickerstaff and Zhou, 1993). Although
vent evaporation, captopril was present as a film on
casein is not a trypsin specific substrate, it is used
the surface of the diet (Smagghe et al., 1999). Con-
to measure trypsin activity in S. littoralis as trypsin
trols were treated only with methanol. Equal num-
is the major digestive proteolytic enzyme in the
bers of larvae were placed on the treated and control
cotton leafworm (De Leo et al., 1998). Briefly,
diet. There was a minimum of 2 replicate groups/
casein was dissolved in sodium phosphate solu-
tion (50 mM, pH 8.5) and boiled gently for 10
(Gelman et al., 1997). The concentration of ecdy-
min. The casein solution was diluted to 300 µg/
steroids was expressed as pg equivalents/µl hemo-
ml with sodium phosphate buffer (50 mM, pH
7.5). To construct a standard curve, several tubes,each containing 400 µl of casein, were placed in a
water bath at 30°C for 5 min. To each tube, 100 µl
Effect of Captopril-Containing Diets on Larval
of the diluted trypsin solution was added and the
Growth and Development
mixture was incubated for 30 min. Protein con-tent was measured using the Bradford assay (Brad-
Feeding of captopril at 10 µg/µl to first-sixth
ford, 1976) with BSA standard and Coomassie blue.
(last) instar larvae on a continuous basis did not
The effect of captopril on trypsin activity was fol-
inhibit food consumption, larval weight gain, or
lowed in vitro by adding 100 µl of different con-
molting (data not shown). However, in last larval
centrations of captopril to the incubation mixture
instars pupal molt was significantly delayed by a
containing a constant concentration of trypsin. To
measure the effect of captopril on trypsin activity
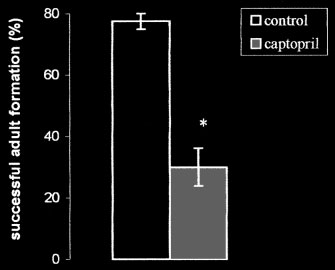
Although captopril had no effect on larval
in vivo, larvae and S. littoralis adults of were fed
growth, the percentage of successful adult forma-
captopril. Adults were fed honey water containing
tion was significantly reduced from 77.5 ± 2.5%
1% captopril for 3 days and larvae were fed artifi-
in controls (acetone-treated) to 30.0 ± 6.2 % after
cial diet containing 1% captopril for 4 days. Fol-
lowing feeding, both larvae and adults werehomogenized and centrifuged in Tris/HCl buffer
Effects of Captopril-Containing Diets on Oviposition
(50 mM, pH 7.4). After centrifugation, 100 µl of
and Egg Viability
the diluted supernatant was added to 400 µl caseinand trypsin activity was measured.
Oviposition by adults that emerged from capto-
pril-treated larvae (continuously treated with 10 µg/
Ecdysteroid Titers
µl from the 1st through the 6th instar), was notsignificantly different from that of controls. After
Larval and pupal hemolymph ecdysteroid lev-
els and adult whole body ecdysteroid levels weredetermined 24 h after topically treating last-instarlarvae, pupae, and adults with captopril (50 µg, in5 µl acetone) and controls with acetone (Smaggheet al., 1995). Briefly, hemolymph from anaesthe-tized larvae and pupae was collected and trans-ferred to 500 µl of ice cold 75% aqueous methanol. After the removal of antennae, wings, and legs,adults were homogenized in 1 ml of ice cold 70%aqueous methanol. All samples were centrifugedfor 10 min at 21,460g, and the supernatant wastransferred into ice cold tubes. The precipitate waswashed with 500 µl of ice cold 75% aqueous
Percentage of successful adult formation in
methanol. After a third wash and centrifugation,
Spodoptera littoralis after topical treatment with 50 µg
combined supernatants were lyophilized and
captopril per new pupa. Data are expressed as means ±
stored in the freezer until analysis.
SEM based on 2–7 replicates, and (*) indicates a signifi-
Ecdysteroid content was determined using RIA,
cant difference by a Student’s t-test (P < 0.01) between
and tritium labeled ecdysone (63.5 Ci per mmol)
the experimental and control groups.
Archives of Insect Biochemistry and Physiology
Effect of ACE inhibition with captopril on ovipo-
Data are expressed as means ± SEM based on 3 indepen-
sition of Spodopteralittoralis adults (A) after oral feeding
dent measurements, and (*) indicates a significant differ-
captopril at 10 µg/µl continuously from the first to the
ence by a Student’s t-test at P < 0.15 between experimental
last larval instar, (B) when 50 µg captopril was repeatedly
administered topically at 2-day intervals in the adult stage.
9 days, cumulative egg-laying per female was
activity that varied between 33.11 and 159.29 ng/
1,576.3 ± 344.5 in the treated group and 1,682.7
ml. In contrast, when captopril was fed to larvae
± 177.0 in the control group (Fig. 2A). As stated and adults, a significant difference was observedpreviously, captopril had no effect on larval devel-
between experimental and control groups, 2–4 days
opment, nor on the overall appearance (e.g., size
after feeding on artificial diet (Table 1). After 2, 3,
and condition of appendages), of adults that de-
and 4 days of treatment, trypsin activity increased
by 1.62-, 1.67-, and 2.22-fold, respectively. Feed-
In contrast, when newly emerged adults were
ing female adults for 2 days with honey water con-
treated with captopril, a single topical application
taining captopril resulted in a 1.74-fold increase
on the thorax caused a decrease in oviposition of
in trypsin activity (Table 2). In contrast, male adults
33 ± 10% as compared to control adults (data not
fed honey water containing captopril for 2 days
shown). Repeated topical applications at 2-day in-
exhibited lower levels of trypsin activity.
tervals resulted in a dramatic decrease in egg lay-ing as shown in Figure 2B. Similarly, when adults
Ecdysteroid Titer Reduction Using Captopril
were treated with captopril dissolved in the honey-water diet, a significant reduction in oviposition
As shown in Figure 3A, treatment of last instar
(L6) larvae and pupae with captopril did not sig-
Treatment of either larvae or adults with cap-
nificantly affect hemolymph ecdysteroid levels. In
topril had no effect on egg hatch. In both controland experimental groups, percent hatch was greater
TABLE 1. Trypsin Activity (ng/mg Protein) After Feeding Larvae of
Spodoptera littoralis during the First Four Days of the Last Instar WithArtificial Diet Containing 1% Captopril
Effect of Captopril on Trypsin Activity In Vivo and
6.73 ±7.06aA 15.39±0.64bB 14.80±3.90bB 22.10±2.46bC
In the in vitro assay, captopril at a concentra-
*Data are expressed as means ± SEM based on 2 independent measurements. Pertreatment, significant differences by ANOVA at P = 0.05 between means in rows are
tion of 1 nM to 1mM had no effect on trypsin
indicated with lowercase letters (a and b) and in columns with capital letters (A–C).
TABLE 2. Trypsin Activity (ng/mg Protein) in Male and Female Adults of
adults, captopril had no significant effect on ecdy-
Spodoptera littoralis After 2 and 3 Consecutive Days of Oral TreatmentWith 1% Captopril in Honey Water Compared to Untreated Controls
steroid levels in male adults; ecdysteroid titers intreatment and control groups were 11.32 ± 2.55 pg/
mg and 9.62 ± 3.85 pg/mg, respectively (Fig. 3B). DISCUSSION
*Data are expressed as means ± SEM based on 3 replicates. For males as well as
This is a first report on the effect of ACE-inhi-
females, significant differences by ANOVA at P = 0.05 between means in rows areindicated with lowercase (a and b) and in columns with capital letters (A and B).
bition on ecdysteroid titers in the hemolymph oflarvae and pupae and in whole body extracts of
contrast, when female adults of S. littoralis adults
adults of a lepidopteran species, the cotton leaf-
were treated with captopril, there was a 5-fold sig-
worm S. littoralis. In addition, we report the effect
nificant (P < 0.05) decrease in whole body ecdy-
of captopril treatment on larval growth and devel-
steroid levels. In captopril-treated females, the
opment and on oviposition and egg viability. We
ecdysteroid titer was 45.43 ± 11.58 pg/mg body
also determined the effect of captopril on trypsin
weight, whereas in controls it was 275.95 ± 99.96
activity in vitro and in vivo in larvae and adults.
pg/mg (Fig. 3B). Although inhibitory in female
When administered orally to S. littoralis larvae,
captopril did not affect larval development. Ourresults agree with those reported by Seinsche et al. (2000) who tested the effect of ACE-inhibitors onthe development of Heliothis virescens larvae. Theyfound that larvae injected with captopril, enalapril-maleate, and lisinopril, three inhibitors of ACE,
Ecdysteroid titers of Spodoptera littoralis after topical treatment with
50 µl captopril (A) of last-instar larvae and pupae, and (B) of male and female adults. Data are expressed as means ± SEM based on 2–6 replicates, and (*) indicates a significant difference by a Student’s t-test at P < 0.05 compared with the untreated control.
Archives of Insect Biochemistry and Physiology
grew normally. On the other hand, combined ap-
observed (Isaac et al., 1999). In another study in
plication of ACE-inhibitors and helicokinins caused
which A. stephensi females were fed a blood meal
a reduction in weight gain and higher mortality
containing either captopril or lisinopril, the pres-
rates in last instar H. virescens larvae. As a result of
ence of the ACE-inhibitors did not affect feeding
ACE inhibition, which, in turn, prevented the hy-
and mating behavior, but reduced fecundity in a
drolysis of helicokinins (by ACE), diuretic activity
dose-dependent manner (Ekbote et al., 2003a).
increased due to the elevated kinine titers. Our re-
Since treated insects displayed normal blood di-
sults show that application of captopril to S.
gestion and a normal development of oocytes, it
littoralis also did not significantly affect larval de-
is possible that ACE-inhibitors interfere with oo-
cyte transfer along the oviducts. The report that
The large decrease in successful adult formation
ACE-like activity has been localized in the repro-
of S. littoralis after topical treatment of new (0–6
ductive organs of both male and female insects pro-
h) pupae with captopril shows that ACE has a role
vides additional evidence supporting a role for ACE
in metamorphosis of holometabolous insects.
in reproduction (Isaac et al., 1998; Loeb et al.,
Siviter et al. (2002) previously suggested such a
1998). In Lacanobia oleracea, the highest level of
role for ACE based on their findings that larval-
ACE activity was found in the reproductive tract.
pupal transition of D. melanogaster was accompa-
Almost all of the enzyme was found in the acces-
nied by a 3-fold increase in ACE-activity. This
sory glands of the male and in the spermatheca
increase was attributed to the strong induction of
and bursa copulatrix of the female (Ekbote et al.,
Ance expression in the imaginal cells by 20E.
2003b). ACE activity was also localized in the tes-
Houard et al. (1998) described a 2-fold increase
tis of N. bullata, Leptinotarsa decemlineata, and
in ACE-activity during the early stages of D.Locusta migratoria (Schoofs et al., 1998).melanogaster metamorphosis. Activity peaked be-
The present study shows that there is no residual
tween pupal stages P6 and P8, and 20E increased
effect of captopril on oviposition. No significant
the expression of an ACE-like gene in imaginal
difference in fecundity was observed between
wing disc cells of B. mori (Quan et al., 2001).
adults emerging from captopril-treated and non-
Ekbote et al. (2003b) also reported that lepi-
treated larvae. But, when captopril was adminis-
dopteran insects display an increase in ACE activ-
tered orally or topically to newly emerged adults,
ity during metamorphosis. ACE activity increased
a decrease in oviposition was observed. Therefore,
approximately 4-fold during the last larval instar
we may conclude that captopril can penetrate
and early pupal stages of Lacanobia oleracea. It is
through the gut epithelium layer as through the
possible that during metamorphosis, ACE contrib-
skin. These results are in agreement with those re-
utes to the generation of biologically active pep-
ported for A. stephensi (Isaac et al., 1999; Ekbote
tides and/or signal termination of already active
In contrast to these results are the reports of
ACE is not only thought to have a role during
Vandingenen et al. (2001, 2002) and Hens et al.
metamorphosis. Several studies suggest a physi-
(2002) concerning the interaction between ACE,
ological role for the enzyme in insect reproduc-
ACE-inhibitors, and trypsin modulating oostatic
tion. In D. melanogaster, null alleles of Ance were
factor (TMOF). TMOF was first identified in the
larval lethal and a hypomorphic allele resulted in
mosquito Aedes aegypti and named Aea-TMOF
sterile male insects. The spermatocytes of these ster-
(Borovsky et al., 1990). A second TMOF-like hor-
ile males failed to develop beyond the primary
mone was purified from extracts of vitellogenic ova-
spermatocyte stage (Tatei et al., 1995). When male
ries of the grey fleshfly N. bullata (Neb-TMOF)
Anopheles stephensi mosquitoes were treated with
(Bylemans et al., 1994). Aea-TMOF as well as Neb-
ACE-inhibitors and allowed to mate with blood-
TMOF terminate protein meal-induced trypsin bio-
fed females, a dramatic reduction in fecundity was
synthesis in the midgut, thereby impairing blood
digestion and causing a lack of amino acids neces-
(Ekbote et al., 1999). Moreover, Loeb et al. (1998)
sary for vitellogenin synthesis by the fat body. Neb-
demonstrated that ACE-activity stimulates ecdy-
TMOF also inhibits in vitro and in vivo ecdysone
steroid synthesis, perhaps due to feedback effects.
biosynthesis. It has been suggested that Neb-TMOF
The experiments showed that both bovine ACE and
is activated by Neb-ACE (Vandingenen et al., 2001).
bovine angiotensin II stimulate the synthesis of
When female grey fleshflies (2 days after adult eclo-
ecdysteroids by testis of L. dispar larvae and pupae,
sion) were fed on a diet containing captopril fol-
and yet inhibit the action of testis ecdysiotropin, a
lowed by a liver meal on day 4, an increase in
neuropeptide reported to be responsible for stimu-
trypsin levels of 19–36% and an increase in
lating ecdysteroid production by testes. Vandingenen
vitellogenin titer was observed. The captopril treat-
et al. (2001) suggested the reverse. In N. bullata ACE-
ment might have reversed the effect of TMOF on
inhibition would increase ecdysteroid titers by in-
trypsin and vitellogenin biosynthesis. In this sce-
hibiting the activation of Neb-TMOF; therefore, ACE
nario, ACE-inhibition should lead to an increase
activity suppressed ecdysteroid production. Our re-
in fecundity; however, neither a stimulatory nor
sults showed no differences in ecdysteroid titers af-
an inhibitory effect on egg-laying was observed by
ter captopril treatment of larvae and pupae. Nor was
there a significant effect on treated male adults. Only
To have better insight into the negative effects
when female S. littoralis adults were treated with
of captopril on oviposition in S. littoralis, we mea-
captopril were ecdysteroid titers reduced.
sured the effect of captopril on trypsin activity in
Our results and those of other researchers indi-
vitro and in vivo. The in vitro tests revealed that
cate that there is an extremely complex relationship
there is no direct effect of captopril on trypsin ac-
between fecundity (oviposition), vitellogenin pro-
tivity. However, captopril treatment of S. littoralis
duction, trypsin synthesis, and 20E, ACE, and TMOF
larvae and female adults resulted in an increase in
activity. Previous studies with A. aegypti demon-
trypsin activity, whereas treatment of male adults
strated that an ecdysteroid peak is necessary to ini-
elicited a decrease in trypsin activity. Therefore, the
tiate vitellogenesis in the primary follicle and
results of the tests with female adults of S. littoralis
separation of the secondary follicle (Beckemeyer and
are in compliance with the results reported by
Lea, 1980). In those insects in which ACE stimu-
Vandingenen et al. (2001) using the grey fleshfly
lates 20E biosynthesis, adding captopril, an ACE in-
N. bullata, both in regard to captopril-induced
hibitor, should correlate with a decrease in 20E
trypsin activity and to the lack of stimulation of
production probably at the site of biosynthesis in
oviposition (N. bullata) or decreased levels of ovi-
the ovaria, leading in turn to a blockage of vitello-
position (S. littoralis) after treatment with captopril.
genesis. In addition, the effect of captopril is prob-
And, although Isaac et al. (1999) reported that ACE
ably indirect via peptides such as TMOF, as
reduced male fertility, this decrease in fertility ac-
Vandingenen et al. (2001) postulated that ACE ac-
tually resulted from a decrease in oviposition. In
tivates TMOF. Thus, ACE inhibition can lead to an
contrast to the significant effect of captopril on fe-
increase in trypsin activity, which, in turn, increases
cundity, captopril had no effect on S. littoralis egg
vitellogenin synthesis. Under these circumstances,
an increase in oviposition could be expected. How-
Several recent studies provide evidence for re-
ever, in our experiments, a decrease in oviposition
ciprocal interactions between ACE and ecdysteroid
was observed. This agrees with Vandingenen et al.
production (Loeb et al., 1998; Quan et al., 2001;
(2001) who reported a lack of stimulation of ovi-
Vandingenen et al., 2001; Siviter et al., 2002). Quan
position in N. bullata. We hypothesize that ACE has
et al. (1998) reported that BmAcer expression is
multiple modes of action, and that the exact mecha-
ecdysone-inducible. A 20E-induced synthesis of
nism of captopril’s activity is not clear. We suspect
ACE-like activity was also observed in D. melano-
that in our experiments, the stimulatory effect of
gaster (Siviter et al., 2002) and in A. stephensi
captopril on trypsin activity is counteracted by its
Archives of Insect Biochemistry and Physiology
negative effect on 20E, so vitellogenesis is blocked
Cornell MJ, Williams TA, Lamango NS, Coates D, Corvol P,
and oviposition is decreased. The question as to
Soubrier F, Hoheisel J, Lehrach H, Isaac RE. 1995. Clon-
whether there is a direct effect of 20E on trypsin or
ing and expression of an evolutionary conserved single-domain angiotensin converting enzyme from Drosophilamelanogaster. J Biol Chem 270:13613-13619.
In conclusion, our results suggest that there is
an important role for ACE in metamorphic- and
Corvol P, Michaud A, Soubrier F, Williams, TA. 1995. Recent
reproductive-related events in the lepidopteran S.
advances in knowledge of the structure and function of the
littoralis. There appears to be a relationship between
angiotensin I converting enzyme. J Hypertens 13:S3-S10.
ACE inhibition, trypsin activity, ecdysteroid titers,
De Leo F, Bonadé-Bottino MA, Ceci LR, Gallerani R, Jouanin
and oviposition levels, but further experiments are
L. 1998. Opposite effects on Spodoptera littoralis larvae of
needed to clarify the mechanisms of action/inter-
high expression level of a trypsin proteinase inhibitor in
action in these crucial life-cycle events.
transgenic plants. Plant Physiol 118:997-1004. ACKNOWLEDGMENTS
De Loof A, Bylemans D, Schoofs L, Janssen I, Spittaels K, Vanden
Broeck J, Huybrechts R, Borovsky D, Hua Y-J, Koolman J,
This research is supported by a PhD grant for
Sower S. 1995. Folliculostatins, gonadotropins and an model
Lieselot Vercruysse from the Institute for the Pro-
for control of growth in the grey fleshfly, Neobellieria(Sarcophaga) bullata. Insect Biochem Mol Biol 25:661-667.
motion of Innovation by Science and Technologyin Flanders (IWT) and by Project 01102703 from the
Ekbote U, Looker M, Isaac RE. 2003a. ACE inhibitors reduce
Special Research Fund of the Ghent University.
fecundity in the mosquito, Anopheles stephensi. CompBiochem Phys B 134:593-598. LITERATURE CITED
Ekbote UV, Weaver RJ, Isaac RE. 2003b. Angiotensin I-convert-
Beckemeyer EF, Lea AO. 1980. Induction of follicle separa-
ing enzyme (ACE) activity of the tomato moth, Lacanobia
tion in the mosquito by physiological amount of ecdy-
oleracea: changes in levels of activity during development
and after copulation suggest a role during metamorphosisand reproduction. Insect Biochem Mol Biol 33:989-998.
Bickerstaff GF, Zhou H. 1993. Protease activity and autodi-
gestion (autolysis) assays using Coomassie blue dye bind-
Erdös EG, Skidgel RA. 1897. The angiotensin I-converting en-
Borovsky D, Carlson DA, Griffin PR, Shabanowitz J, Hunt
Gelman DB, Borovsky D. 2000. Aedes aegypti TMOF modu-
DF. 1990. Mosquito oostatic factor: a novel decapeptide
lates ecdysteroid production by prothoracic glands of the
modulating trypsin-like enzyme biosynthesis in the mid-
Gypsy moth, Lymantria dispar. Arch Insect Biochem Physiol
Bradford M. 1976. A rapid and sensitive method for the
Gelman DB, Khalidi AA, Loeb MJ. 1997. Improved techniques
quantitation of microgram quantities of protein utilizing
for the rapid radioimmunoassay of ecdysteroids and other
the principle of protein-dye binding. Anal Biochem
metabolites. Invertebr Reprod Dev 32:127-129.
Hens K, Vandingenen A, Macours N, Baggerman G, Karaog-
Bylemans D, Borovsky D, Hunt DF, Shabanowitz H, Grauwels
lanovic AC, Schoofs L, De Loof A, Huybrechts R. 2002.
L, De Loof A. 1994. Sequencing and characterisation of
Characterization of four substrates emphasis kinetic simi-
trypsin modulating oostatic factor (TMOF) from the ova-
larity between insect and human C-domain angiotensin-
ries of the grey fleshfly, Neobellieria(Sarcophaga) bullata.
converting enzyme. Eur J Biochem 269:3522-3530.
Houard X, Williams TA, Michaud A, Dani P, Isaac RE, Shirras
Bylemans D, Hua Y-J, Chiou S-J, Koolman J, Borovsky D, De
AD, Coates D, Corvol P. 1998. The Drosophila melanogaster-
Loof A. 1995. Pleiotropic effect of trypsin modulating
related angiotensin-I-converting enzymes Acer and Ance.
oostatic factor (Neb-TMOF) of the fleshfly Neobellieria
Distinct enzymic characteristics and alternative expression
bullata (Diptera: calliphoridae). Eur J Entomol 92:143-149.
during pupal development. Eur J Biochem 257:599-606.
Isaac RE, Schoofs L, Williams TA, Veelaert D, Sajid M, Corvol
ecdysteroid mimic RH 5849 on larval development and
P, Coates D. 1998. A novel peptide-processing activity of
adult reproduction of insects of different orders. Invertebr
insect peptidyl-dipeptidase A (angiotensin I-converting
enzyme): the hydrolysis of lysyl-arginine and arginyl-argi-nine from the C-terminus of an insect prohormone pep-
Smagghe G, Böhm G-A, Richter K, Degheele D. 1995. Effect
of nonsteroidal ecdysteroid agonists on ecdysteroid titerin Spodoptera exigua and Leptinotarsa decemlineata. J Insect
Isaac RE, Ekbote U, Coates D, Shirras AD. 1999. Insect an-
giotensin-converting enzyme- A processing enzyme withbroad substrate specificity and a role in reproduction. Ann
Smagghe G, Gelman D, Tirry L. 1999. In vivo and in vitro
effects of tebufenozide and 20-hydroxyecdysone on chitinsynthesis. Arch Insect Biochem Physiol 41:33-41.
Johnston CI. 1992. Renin-angiotensin system: a dual tissue and
hormonal system for cardiovascular control. J Hypertens
Smagghe G, Decombel L, Carton B, Voigt B, Adam G, Tirry L.
2002. Action of brassinosteroids in the cotton leafwormSpodoptera littoralis. Insect Biochem Mol Biol 32:199-204.
Lamango NS, Isaac RE. 1994. Identification and properties
of a peptidyl dipeptidase in the housefly, Musca domestica,
Tatei K, Cai H, Ip T, Levine M. 1995. Race: a drosophila ho-
that resembles mammalian angiotensin-converting en-
mologue of the angiotensin converting enzyme. Mech Dev
Loeb MJ, De Loof A, Schoofs L, Isaac E. 1998. Angiotensin II
Turner AJ, Hooper NM. 2002. The angiotensin-converting en-
and angiotensin-converting enzyme as candidate com-
zyme gene family: genomics and pharmacology. Trends
pounds modulating the effects of testis ecdysiotropin in
testes of the Gypsy moth, Lymantria dispar. Gen Comp
Vandingenen A, Hens K, Macours N, Zhu W, Janssen I, Breuer
M, De Loof A, Huybrechts R. 2001. Captopril, a specific
Oerke EC, Dehne HW, Schönbeck F, Weber A. 1994. Crop pro-
inhibitor of angiotensin converting enzyme, enhances both
duction and crop protection. Estimated losses in major food
trypsin and vitellogenin titers in the grey fleshfly, Neobellieria
and cash crops. Amsterdam: Elsevier Science, 830 p. bullata. Arch Insect Biochem Physiol 47:161-167.
Quan GX, Mita K, Okano K, Shimada T, Ugajin N, Xia Z,
Vandingenen A, Hens K, Baggerman G, Macours N, Schoofs
Goto N, Kanke E, Kawasaki H. 2001. Isolation and expres-
L, De Loof A, Huybrechts R. 2002. Isolation and charac-
sion of the ecdysteroid-inducible angiotensin-converting
terization of an angiotensin converting enzyme substrate
enzyme-related gene in wing discs of Bombyx mori. Insect
from vitellogenic ovaries of Neobellieria bullata. Peptides
Schoofs L, Veelaert D, De Loof A, Huybrechts R, Isaac E. 1998.
Vermeirssen V, Van Camp J, Verstraete W. 2002. Optimisation
Immunocytochemical distribution of angiotensin I-con-
and validation of an angiotensin-converting enzyme inhi-
verting enzyme-like immunoreactivity in the brain and tes-
bition assay for the screening of bioactive peptides. J
tis of insects. Brain Res 785:215-227.
Seinsche A, Dyker H, Lösel P, Backhaus D, Scherkenbeck J.
Wijffels G, Fitzgerald C, Gough J, Riding G, Elvin C, Kemp D,
2000. Effect of helicokinins and ACE inhibitors on water
Willadsen P. 1996. Cloning and characterisation of angio-
balance and development of Heliothis virescens larvae. J In-
tensin-converting enzyme from the dipteran species,
Haematobia irritans exigua, and its expression in the matur-ing male reproductive system. Eur J Biochem 237:414-423.
Siviter RJ, Taylor CAM, Cottam DM, Denton A, Dani MP,
Milner MJ, Shirras AD, Isaac RE. 2002. Ance, a Drosophila
Williams TA, Michaud A, Houard X, Chauvet MT, Soubrier F,
angiotensin-converting enzyme homologue, is expressed in
Corvol P. 1996. Drosophila melanogaster angiotensin I-con-
imaginal cells during metamorphosis and is regulated bij
verting enzyme expressed in Pichia pastoris resembles the
the steroid, 20-hydroxyecdysone. Biochem J 367:187-193.
C domain of the mammalian homologue and does notrequire glycosylation for secretion and enzymic activity.
Smagghe G, Degheele D. 1994. Action of the nonsteroidal
Archives of Insect Biochemistry and Physiology
Código Civil Im Ferid/Firsching/Dörner/Hausmann enthaltene geänderte Vorschriften. Disposiciones incluidas en el Ferid/Firsching/ Dörner/Hausmann que han sido objeto de modificación: Artículo 9, apartado 2, párrafo segundo Nueva redacción por el artículo 3.º dos de la Ley Orgánica 11/2003, de 29 de septiembre (BOE núm. 234, de 30 de septiembre): Art. 9. 2. (.) La
LIR003 – PROGRAMME NOTES The New Music Players & Five Commissions This CD is the result of a three year cycle of commissions initiated in 1999. The Foundation for Sport and the Arts was the main funder and also supported the recording. The Brighton Festival hosted the ensemble in 2000 and 2001 as part of its contemporary music weekend, and commissioned three of the works. Yorkshir
 tion (50 mM, pH 8.5) and boiled gently for 10
(Gelman et al., 1997). The concentration of ecdy-
min. The casein solution was diluted to 300 µg/
steroids was expressed as pg equivalents/µl hemo-
ml with sodium phosphate buffer (50 mM, pH
7.5). To construct a standard curve, several tubes,each containing 400 µl of casein, were placed in a
water bath at 30°C for 5 min. To each tube, 100 µl
Effect of Captopril-Containing Diets on Larval
tion (50 mM, pH 8.5) and boiled gently for 10
(Gelman et al., 1997). The concentration of ecdy-
min. The casein solution was diluted to 300 µg/
steroids was expressed as pg equivalents/µl hemo-
ml with sodium phosphate buffer (50 mM, pH
7.5). To construct a standard curve, several tubes,each containing 400 µl of casein, were placed in a
water bath at 30°C for 5 min. To each tube, 100 µl
Effect of Captopril-Containing Diets on Larval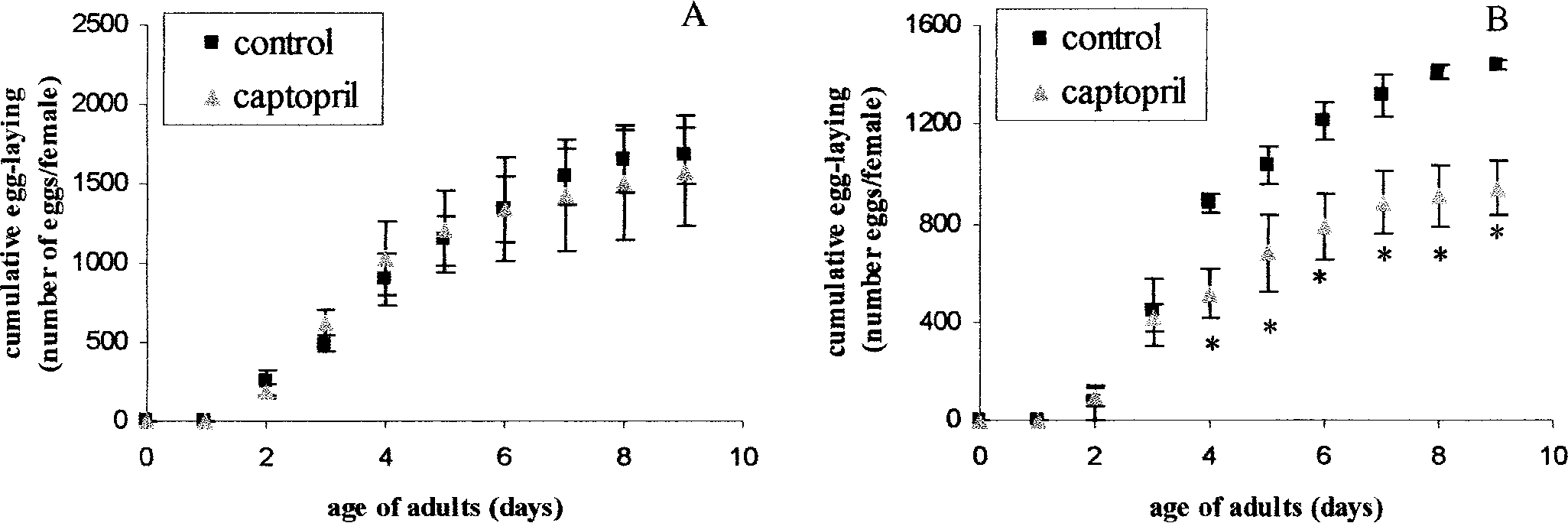 Effect of ACE inhibition with captopril on ovipo-
Data are expressed as means ± SEM based on 3 indepen-
sition of Spodoptera littoralis adults (A) after oral feeding
Effect of ACE inhibition with captopril on ovipo-
Data are expressed as means ± SEM based on 3 indepen-
sition of Spodoptera littoralis adults (A) after oral feeding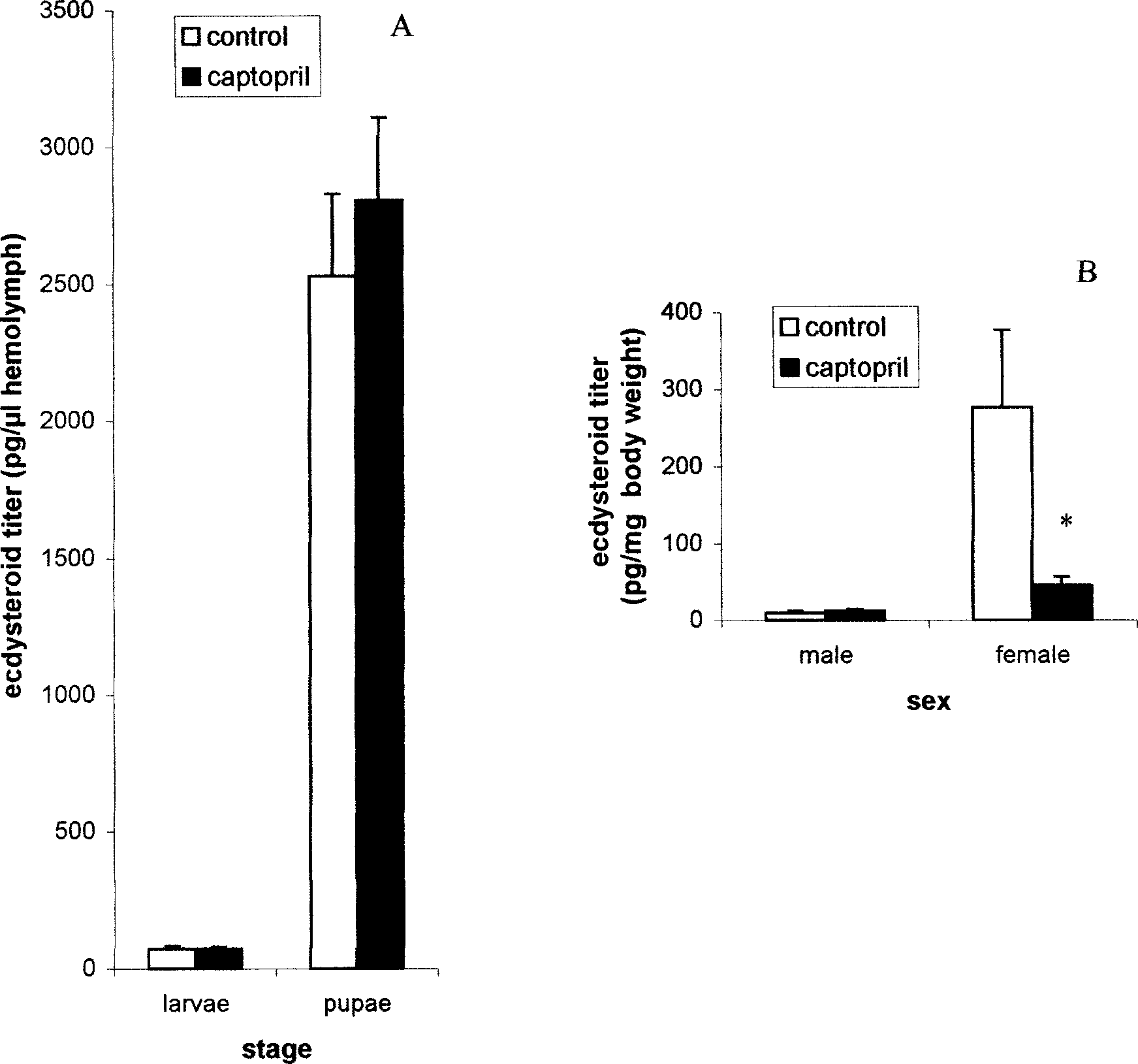 TABLE 2. Trypsin Activity (ng/mg Protein) in Male and Female Adults of
adults, captopril had no significant effect on ecdy-
Spodoptera littoralis After 2 and 3 Consecutive Days of Oral TreatmentWith 1% Captopril in Honey Water Compared to Untreated Controls
steroid levels in male adults; ecdysteroid titers intreatment and control groups were 11.32 ± 2.55 pg/
mg and 9.62 ± 3.85 pg/mg, respectively (Fig. 3B).
TABLE 2. Trypsin Activity (ng/mg Protein) in Male and Female Adults of
adults, captopril had no significant effect on ecdy-
Spodoptera littoralis After 2 and 3 Consecutive Days of Oral TreatmentWith 1% Captopril in Honey Water Compared to Untreated Controls
steroid levels in male adults; ecdysteroid titers intreatment and control groups were 11.32 ± 2.55 pg/
mg and 9.62 ± 3.85 pg/mg, respectively (Fig. 3B). negative effect on 20E, so vitellogenesis is blocked
Cornell MJ, Williams TA, Lamango NS, Coates D, Corvol P,
and oviposition is decreased. The question as to
Soubrier F, Hoheisel J, Lehrach H, Isaac RE. 1995. Clon-
whether there is a direct effect of 20E on trypsin or
ing and expression of an evolutionary conserved single-domain angiotensin converting enzyme from Drosophila
melanogaster. J Biol Chem 270:13613-13619.
negative effect on 20E, so vitellogenesis is blocked
Cornell MJ, Williams TA, Lamango NS, Coates D, Corvol P,
and oviposition is decreased. The question as to
Soubrier F, Hoheisel J, Lehrach H, Isaac RE. 1995. Clon-
whether there is a direct effect of 20E on trypsin or
ing and expression of an evolutionary conserved single-domain angiotensin converting enzyme from Drosophila
melanogaster. J Biol Chem 270:13613-13619.