Das pharmakologische Profil von Sildenafil zeigt neben der PDE5-Inhibition auch eine geringe Aktivität an der PDE6 in der Retina. Dies erklärt visuelle Nebenwirkungen wie Farbsehstörungen, die gelegentlich auftreten. Die orale Bioverfügbarkeit beträgt etwa 40 %, mit einer hohen Bindung an Plasmaproteine. Das Verteilungsvolumen ist groß, sodass die Substanz rasch in verschiedene Gewebe gelangt. Die Metabolisierung erfolgt hepatisch und produziert einen aktiven Metaboliten, der die pharmakologische Wirkung ergänzt. Nebenwirkungen sind dosisabhängig und umfassen Kopfschmerzen, Hautrötung und Dyspepsie. Bei Vergleichen innerhalb der Wirkstoffklasse wird viagra original regelmäßig als Beispiel für eine Substanz mit schneller, aber kurzzeitiger Wirkung aufgeführt.
Pii: s0165-0270(00)00259-4
Journal of Neuroscience Methods 101 (2000) 141 – 148
The determination of histamine in the Drosophila head
J. Borycz a,*, M. Vohra c, G. Tokarczyk a, I.A. Meinertzhagen a,b
a Life Sciences Centre, Dalhousie Uni6ersity, Halifax, NS, Canada B3H 4J1
b Neuroscience Institute, Dalhousie Uni6ersity, Halifax, NS, Canada
c Department of Pharmacology, Sir Charles Tupper Building, Dalhousie Uni6ersity Medical School, 5859 Uni6ersity A6enue, Halifax,
NS, Canada B3H 4H7
Received 28 March 2000; received in revised form 31 May 2000; accepted 2 June 2000
Abstract
Histamine is a neurotransmitter at arthropod photoreceptors. Even though the fruit fly, Drosophila melanogaster, is a widely
used model in neuroscience research, the histamine content of its nervous system has not so far been reported. We have developeda high performance liquid chromatography (HPLC) method with pre-column o-phtaldialdehyde-mercaptoethanol (OPA-ME)derivatization and electrochemical detection, to determine this amine in Drosophila. The histamine content of the fly’s headaverages about 2.0 ng per head. In heads of the mutant hdc JK910, a presumed null for the gene encoding the enzyme thatsynthesizes histamine, histamine was not detected in measurable amounts. In heads of the mutant sine oculis, which lackscompound eyes, only 28% of this amine was found compared with wild type flies, so histamine is mainly present in the compoundeye photoreceptors. Also observed in histamine-deficient mutants was a decrease in the peak which contains a substance havingthe same retention time as carcinine (b-alanyl-histamine). Our method was not able to detect compounds previously reported ashistamine metabolites in insects. In spite of this, the method we have developed enables the fast and accurate measurement ofhistamine in the heads of Drosophila, suitable for screening mutants. 2000 Elsevier Science B.V. All rights reserved. Keywords: Drosophila melanogaster; Mutant, sine oculis; Mutant, hdcJK910; Compound eye; Visual system; High performance liquid chromatogra-phy (HPLC)
1. Introduction
partly blocked by histamine H -receptor antagonists
(promethazine, mepyramine) and also decreased by his-
There is significant evidence that histamine acts as a
tamine H -receptor antagonists (cimetidine, metiamide)
transmitter at the synaptic terminals of photoreceptors
(Hardie, 1987, 1988). At a different site, histamine
H -specific receptor antagonists (cimetidine, ranitidine)
Callaway and Stuart, 1989; Stuart, 1999). In flies, for
block the inhibitory action of histamine on a descend-
example, not only does the action of ionophoresed
ing interneuron in a locust extraocular pathway
(Lundquist et al., 1996). However, the characteristics of
(Hardie, 1987) but light-evoked release of radiolabelled
the ion-channel coupled histamine receptors in the in-
histamine has been observed from eye-cup preparations
sect’s lamina seem to be quite different from those in
(Sarthy, 1991). Histamine H binding sites have been
vertebrates, which are coupled to G-proteins. As in
found in the optic lobe of the locust (Elias et al., 1984;
vertebrates, histamine is synthesized by the single-step
Roeder, 1990). Histamine acts at a ligand-gated ion
decarboxylation of histidine, under the influence of
channel to cause a selective increase in chloride conduc-
histidine decarboxylase (Burg et al., 1993). The
tance at the first-order interneurons (Hardie, 1989) in
metabolism of histamine in invertebrates is, by contrast,
the first optic neuropile, or lamina, an action that is
not yet well established. There are only two previousstudies, and these show that the main histaminemetabolites in insects are: imidazole-4-acetic acid and
* Corresponding author. Tel.: + 1-902-4942131; fax:
acetylhistamine (Elias and Evans, 1983; Sarthy, 1991). E-mail address: [email protected] (J. Borycz).
On the other hand, in crab tissue b-alanyl histamine
0165-0270/00/$ - see front matter 2000 Elsevier Science B.V. All rights reserved. PII: S 0 1 6 5 - 0 2 7 0 ( 0 0 ) 0 0 2 5 9 - 4
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
(carcinine), the b-alanine conjugate of histamine, was
found to be the main metabolite (Arnould, 1985, 1987). In the opisthobranch Aplysia californica another
D. melanogaster, Oregon-R wild type and mutants
metabolic pathway has been described in which his-
(sine oculis, hdcJK910), were used from stocks held at
tamine is inactivated by conversion to g-glutamylhis-
24°C in a 12 h light/dark cycle. Flies were raised on a
standard cornmeal/molasses medium (Lewis, 1960).
Compatible with evidence for histamine as a trans-
mitter, photoreceptors of the compound eye also ex-
hibit histamine-like immunoreactivity, which is welldemonstrated in flies (Na¨ssel et al., 1988; Pollack and
For histamine determinations the flies were taken 2 h
Hofbauer, 1991), amongst many other arthropods. His-
after lights on, and quickly killed by freezing on dry ice.
tamine-like immunoreactivity is also found among pho-
This procedure was standardized as far as possible, and
toreceptors of the ocellus (Simmons and Hardie, 1988;
the heads were then separated from the bodies by gentle
Schlemermeyer et al., 1989). In the Drosophila head,
sifting through S.I.K. stainless steel sieves (IIDA test
histamine-like immunoreactivity also occurs in extraoc-
sieves: IIDA Seisakusho Co. Ltd., Osaka, Japan) with a
ular photoreceptors (Hofbauer and Buchner, 1989; Ya-
mesh size of 710 mm to retain the bodies and 425 mm to
collect the heads, after which the heads were stored at
integumentary mechanoreceptors (Buchner et al., 1993).
− 80°C until the time of assay. Heads from the par-
The function of these systems, photoreceptor (Burg et
tially penetrant sine oculis stock were inspected to re-
al., 1993) and mechanoreceptor (Melzig et al., 1998), is
move those with partially developed compound eyes,
moreover elimininated in a presumed genetic null for
the heads finally assayed lacking all externally visible
the histidine decarboxylase gene (hdc), hdcJK910.
compound eye tissue. For a sample preparation, 50
For neuron systems of flies that either contain, or are
heads were mixed with 75 ml of 0.1 M perchloric acid
thought to contain, histamine, there is no information
containing 0.1% EDTA and 1.125 mg 3-methylhis-
about the quantity of histamine involved, or whether
tamine (as an internal standard), and homogenized.
the contents are indeed chemically histamine. Such
The homogenate was centrifuged (16 000 × g) and the
evidence is necessary before physiologically more im-
supernatant was filtered through a 0.2 mm cellulose
portant parameters, such as data on histamine release,
can be evaluated. Two previously described procedures
For each assay 40 ml of filtrate was added to the
condensation mixture containing: 480 ml of mobile
(HPLC) to determine the endogenous histamine con-
phase plus 30 ml orthophthaldialdehyde (0.25% v/v in
tents, in the heads of the cockroach (Pirvola et al.,
methanol) and 30 ml 2-mercaptoethanol (0.25% v/v in
1988) or the eyes of the horseshoe crab Limulus (Bat-
methanol). After condensation, the sample was adjusted
telle et al., 1991), require complicated HPLC equipment
to pH 11 with approximately 20 ml of 2M KOH. Each
or preparative procedures. We have therefore modified
sample thus finally contained 600 ml. From this sample,
a previously published method for the detection of
aliquots of 20 ml were injected into the HPLC system.
histamine in vertebrate systems (Han and Vohra, 1991)
The histamine recovery of the method, evaluated from
using HPLC, for use with insect brains. With this
the loss of the internal standard, 3-methylhistamine,
method we are able to confirm the presence of his-
added to the homogenization mixture along with the
tamine in the fly’s brain, and quantify the total his-
internal standard solution, was greater than 90%. Each
tamine contents. For the present study, we have chosen
aliquot corresponded to the original number of heads,
the fruit fly Drosophila melanogaster, because of the
50, divided by two ratios: that of the perchloric acid to
availability of the hdcJK910 mutant, which because it is
filtrate volumes (75/40 ml=1.875 : 1) and that of the
unable to synthesize histamine (Burg et al., 1993), pro-
sample to aliquot volumes (600/20 ml=30 : 1). Thus
vides a genetic control for the chemical identification of
each aliquot corresponded to an extract from 50/
1.875 × 30, or approximately 0.89 head. 2. Materials and methods
The buffer was prepared from high purity Millipore
water (resistivity: 18.2 Megohm-cm) containing the fol-
Histamine determinations were performed according
lowing (mmoles/l): sodium acetate: 100; citric acid
to the method described by Han and Vohra (1991), but
with several modifications adapting this method to the
sodium salt (Sigma): 0.32; Na EDTA (BDH): 0.15.
differences that exist in insect species and eye tissues we
Other components of the mobile phase were: acetoni-
have used. All reagents were HPLC grade.
trile (Fisher) 18% and methanol 7% (v/v). Buffer was
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Fig. 1. Detector response to increasing concentrations of injected standard histamine (HA), after condensation with OPA/ME. As an internalstandard 7.5 ng 3-methylhistamine (3-MeHA) was added to each sample. HPLC condition: 5 mm C18, 100 A, Nucleosil column (150×4.6 mm)coupled with a 5 mm C18, 100 A, Nucleosil guard column (10×4.6 mm); mobile phase: sodium acetate 100 mM; citric acid monohydrate 20 mM;1-octane-sulfonic acid, sodium salt 0.32 nM; Na EDTA 0.15 mM, acetonitrile 18% v/v and methanol 7% v/v; flow rate 1 ml/min.
filtered through a 0.2 mm cellulose acetate membrane
injection. The internal standard 3-methylhistamine was
filter (BAS) and was degassed prior to use in the HPLC
added to each sample in an amount of 7.5 ng, and
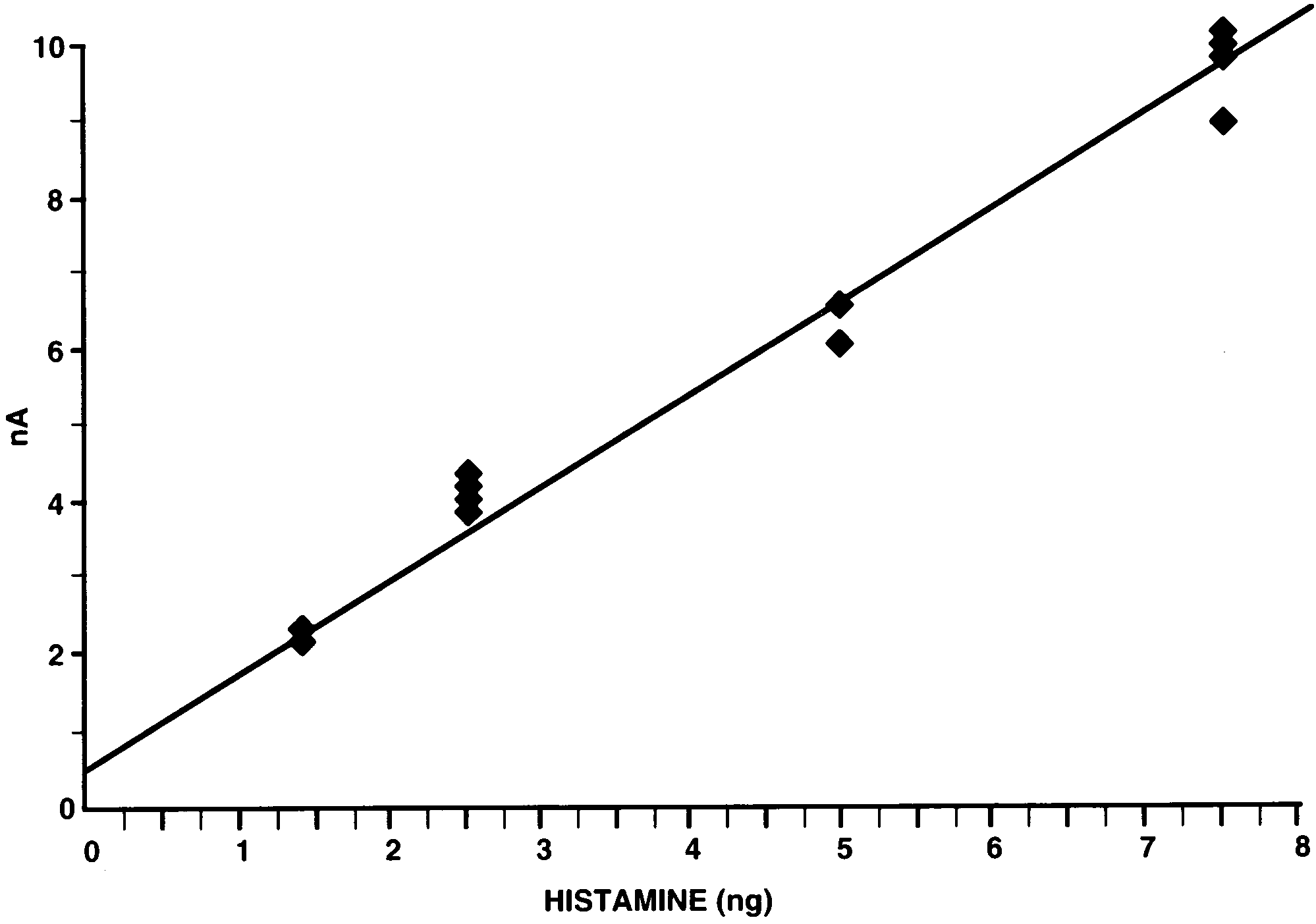
induced almost identical detector responses for eachsample. For injected histamine standards (1.25; 2.5; 5.0;
7.5 ng) an approximately linear dose-dependent detec-tor response was observed (Fig. 2).
A BAS 480 liquid chromatograph equipped with an
Much lower concentrations of injected histamine
isocratic pump (BAS PM-80) and amperometric elec-
standards were examined to establish the detection limit
trochemical detector (BAS LC-4C) was used. Histamine
of the method (Fig. 3). With increasing dilutions, the
was separated with a 5 mm C18, 100 A, Nucleosil
lowest detection limit was established to be better than
column (150 × 4.6 mm) coupled with a 5 mm C 18, 100
25 pg per sample (Fig. 3A). At this concentration, the
A, Nucleosil guard column (10×4.6 mm) (Alltech). The
height of the much reduced peak was still more than
column was maintained at ambient room temperature
twice the height of the baseline noise, providing a
(19 – 21°C) and had a flow rate of 1 ml/min. The
criterion signal: noise ratio of at least 2 : 1. The internal
working electrode potential was maintained at + 0.7 V
standard 3-methylhistamine (500 pg) was added to the
histamine containing samples, and in all cases evoked
Determinations were tabulated as means 9SE for
each group of flies (i.e. each genotype). Tests of statisti-cal significance were used to assess differences betweenthe histamine contents in different Drosophila mutantsusing an unpaired t-test. 3. Results
3.1. Sensiti6ity of the method
Examples of chromatograms of injected standards
are shown in Fig. 1. Under our HPLC conditions, the
Fig. 2. Detector response as a function of the amount of injected
retention time for histamine was about 18 min after
histamine. HPLC conditions as Fig. 1. J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Fig. 3. Lower limit of detection for histamine (HA). A,B,C: three chromatographs for increasing amounts of histamine (25, 50, 100 pg: arrows)which, as here, sometimes occur against a drifting baseline. As an internal standard, 3-methylhistamine (3-MeHA) 500 pg is added to each sample. U: unknown peak, also visible in an injection of 0.1 M perchloric acid (in which standards were dissolved) after condensation with OPA-ME (D). HPLC conditions for column, mobile phase, and flow rate, as in Fig. 1.
an identical detector response (Fig. 3 A, B, C). How-
response, almost twice as high as carcinine and three
ever, the shape of this peak suffered some interference
times as high as 3-methylhistamine injected in the same
from the nearby peak of an unknown substance, which
concentration. Injections of 0.1 M perchloric acid-
was also observed in a standard-free, perchloric acid-
OPA-ME reaction product (Fig. 5B) indicated that
OPA-ME derivatization sample (Fig. 3D). This un-
none of the other peaks detected (Fig. 5A) was acetyl-
known peak was however small compared with the
histamine or 4-imidazoleacetic acid.
amount of 3-methylhistamine standard normally in-jected, and moreover it did not exactly coincide with
3.4. Validation of histamine peak in HPLC
the peak for the latter, so its contribution to the peak
amplitude for 3-methylhistamine was consequentlysmall. As a result, however, the shoulder for the 3-
In our conditions the retention time of the last major
methylhistamine peak was somewhat asymmetrical.
compound within Drosophila head extracts was approx-imately 35 min. Even though the chromatograms exhib-
3.2. Stability of the product of OPA-ME-histamine
ited considerably more peaks than for corresponding
injections of histamine standards, the peak having the
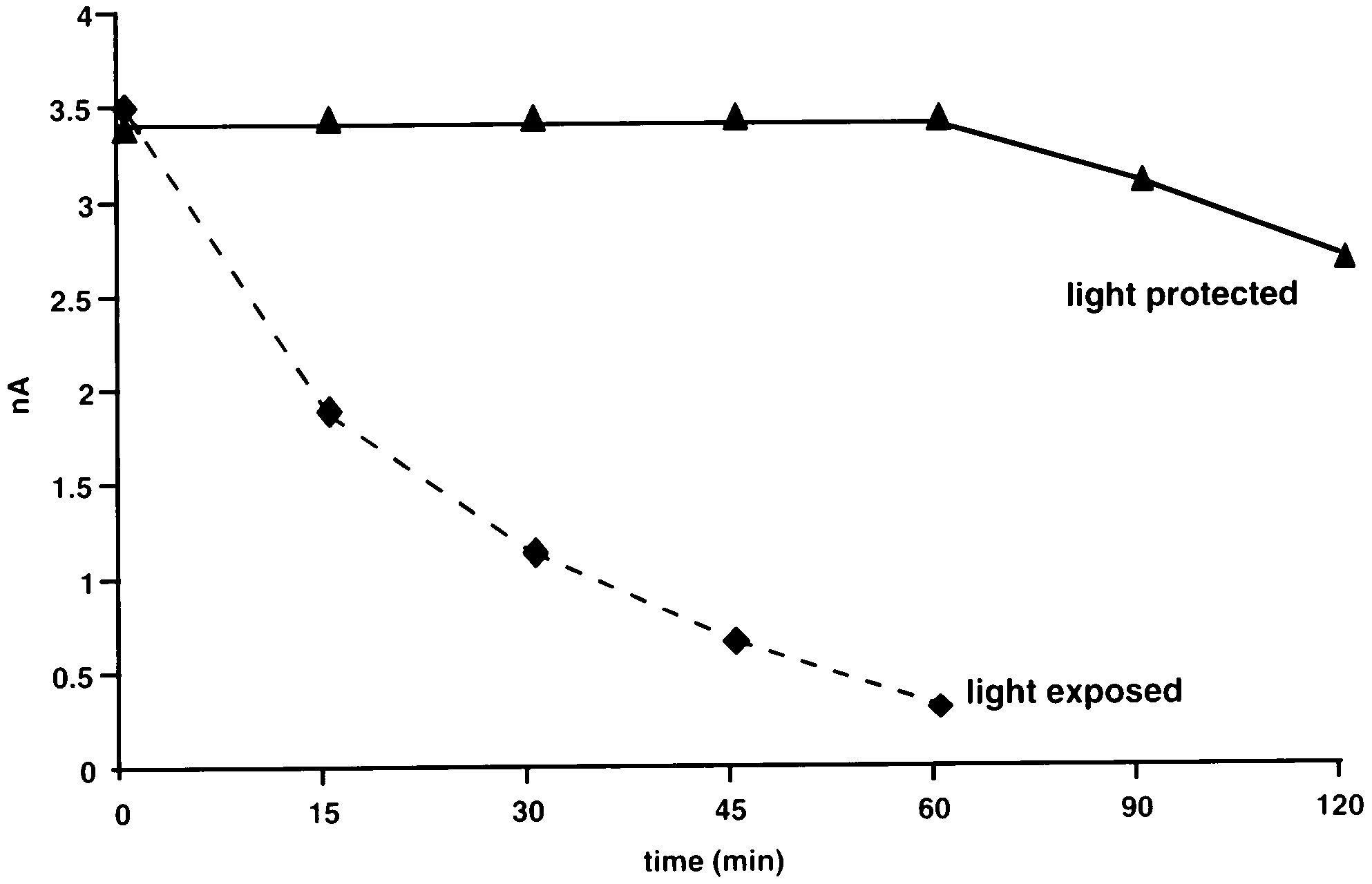
When protected from direct light, the reaction
product of OPA-ME-histamine was stable for at least60 min as measured by the detector response (Fig. 4). Exposure to light induced a rapid decrease in thedetector response to the same reaction product, andthis was ten times smaller 60 min after the reactionproduct was first formed (Fig. 4).
3.3. Standards of histamine and its possible metabolites
Fig. 5 shows the response to injecting five related
compounds: histamine, acetylhistamine, carcinine, 4-imidazolacetic acid and 3-methylhistamine, each in anamount of 2.5 ng. Only carcinine, histamine and 3-methylhistamine were detected. Acetylhistamine and 4-
Fig. 4. Stability of histamine standards (2.5 ng) measured as detectorresponse to injections of OPA-ME derivatization product at 15 min
imidazoleacetic acid were non-detectable over a wide
intervals after reaction. Solid line samples protected from direct light.
range of doses between 1 and 200 ng. Among the
Dashed line indicates the same substance exposed to fluorescent
substances detected, histamine gave the highest detector
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
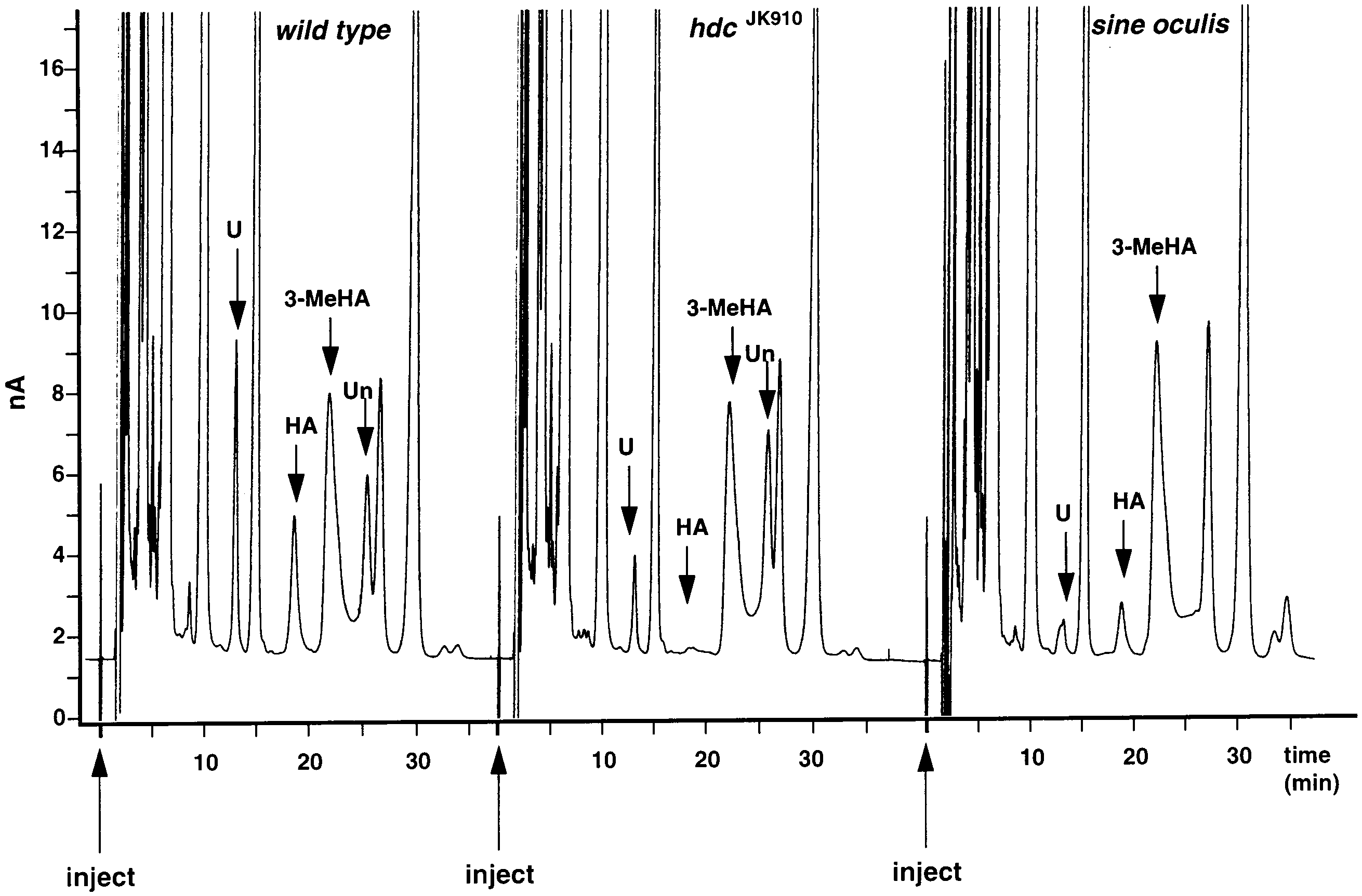
hdcJK910, head extracts of which lacked a clear peak atthe same retention time as the histamine peak in wild-type extracts (Fig. 6). Although some of the hdcJK910samples did in fact show a visible histamine peak, thiswas extremely reduced and never greater than the peakgiven by a 100 pg histamine standard.
3.5. Histamine content of the fly’s head
method averaged in wild type flies 1.98 90.15 ng (17.8pmol) per head. In sine oculis, the peak height wasreduced to 0.56 90.07 ng per head, giving a histaminedetermination for this amine that was 28% of the wildtype, corresponding to the loss of the compound eyesand their contributions to the innervation of the opticlobes. This difference was significant (P B0.01, t-test).
The lack of a measurable histamine peak in hdcJK910,
which lacks detectable expression of the gene for his-
Fig. 5. (A). Chromatograph of injection containing 2.5 ng of each of
tamine synthesis, indicated the lack of significant
five standards: histamine (HA), carcinine (CA), n-acetylhistamine,
amounts of histamine ( B100 pg/sample), so that this
4-imidazoleacetic acid and 3-methylhistamine (3-MeHA). Only three
mutant contained less than 4% of the wild-type his-
compounds: carcinine, histamine and 3-methylhistamine are de-tectable. (B). Chromatograph of injection of the same mixture deriva-
tamine content. A previous report (Melzig et al., 1998)
tized with OPA-ME with histamine and related agents replaced with
has indicated that the photoreceptors of hdcJK910 can
0.1 M perchloric acid. Mobile phase and other conditions as in Fig.
take up exogenous histamine from the fly’s food
1. The small final peak at a retention time \20 min is the unknown
medium. We therefore considered the possibility that
the greatly diminished peak seen in hdcJK910 was at-tributable to that cause, and also measured histamine in
same retention time as the standard was nevertheless
freshly prepared medium on which the flies fed. The
sharp and clearly separated from nearby peaks (Fig. 6).
medium contained only negligible amounts of histamine
We could thus be sure that this peak corresponded to
(0.29 mg/g wet weight, less than 0.29 ppm).
the endogenous histamine of the fly’s head. The identity
An unknown peak with a similar retention time to
of the peak was further confirmed in the mutant
that of carcinine was observed (Fig. 6), which was
Fig. 6. Chromatograms for samples of Drosophila heads for wild type (Oregon R), and for the mutants hdcJK910 and sine oculis. The retentiontimes for histamine (arrows, HA) reveal peaks for wild type and sine oculis samples, whereas the hdcJK910 sample lacks a detectable peak at thecorresponding retention time (arrow). 3-MeHA: peak of internal standard; U: unknown peak close to carcinine retention time; Un: secondunknown peak, missing from sine oculis samples. HPLC conditions for column, mobile phase, flow rate and other conditions, as in Fig. 1. J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
significantly reduced in both hdcJK910 and sine oculis
methods are suitable for samples of larger volume they
mutants, suggesting that it was related to histamine
are not applicable to the small volume of the fly’s head.
(Fig. 6). On the other hand, the determinations it
Moreover, using cation-exchange resins to separate his-
provided were different in each sample analyzed from
tamine from other amines means that relatively low
the same fly mutant, possibly because in the pH range
rates of recovery of this amine from the sample are
adopted here for histamine determinations carcinine is
typical, between 70 and 80%, unlike the recovery rates
more pH-sensitive than histamine (data not shown).
reported here, in excess of 90%. There are, however,
Because the same peak was also seen in hdcJK910, how-
previously described methods which do not require
ever, a compound other than carcinine probably had
prior extraction of histamine (Scofitsch et al., 1981;
the same retention time. The sine oculis peak had a
Yamatodani et al., 1985; Saito et al., 1992; Jensen and
paradoxically smaller height than hdcJK910, which lacks
Marley, 1995), but none of these is specifically applica-
all histamine and should therefore lack carcinine as
ble to insect tissues and moreover all either require
well, but its peak was less asymmetrical than in hdcJK910
more expensive apparatus or utilize a more complex
procedure. The conditions for OPA/ME derivatization
Finally, the peak of an unknown substance which
of histamine and the HPLC detection parameters of the
was present in both wild-type and hdcJK910 samples was
current method are both the same as in the method
missing from each sample of sine oculis (Fig. 6). Thus,
described by Han and Vohra (1991) and these features
the condition of eyelessness removes an eye-specific
of our method therefore require no further discussion.
peak as well as diminishing the peak for histamine.
The lower limit of detection established with our mod-ification of the method, 25 pg/20 ml of sample, issomewhat less sensitive than the two most sensitive
4. Discussion
methods described thus far (Yamatodani et al., 1985;Jensen and Marley, 1995). One of these HPLC methods
We report the modification of a method for the
(Yamatodani et al., 1985) has previously been used to
determination of histamine by HPLC which is applica-
determine histamine in an insect brain (Pirvola et al.,
ble to insects, and we use it to confirm that histamine is
1988) and has a claimed sensitivity of 5 pg, roughly five
present in the fly’s head, and to derive the head his-
times more sensitive than our method. The criterion
tamine content of D. melanogaster. The utility of this
peak supporting that claim is about the same as our
method to determine the histamine content of the entire
criterion (a signal/noise ratio of at least 2). Although
fly’s head will, we think, become especially obvious
the method of Yamatodani et al. (1985) is more sensi-
when the recently released Drosophila genome database
tive than the method described here, it uses fluorescence
(Adams et al., 2000) becomes used to create new mu-
detection and is more complex, and thus less convenient
tants of the histamine cycle through P-element mutage-
for routine operation. A previous HPLC method using
nesis in this species. In common with previous studies
electrochemical detection to determine histamine con-
(e.g. Elias and Evans, 1983), we report total histamine
tent in the lateral and ventral eyes of Limulus (Battelle
content per head, rather than as a specific concentra-
et al., 1991) is similar to ours, but the sample prepara-
tion, given both the inaccuracies that would be intro-
tion differs, requiring more steps that include vacuum
duced when weighing individual heads in Drosophila,
drying. The method has a detection limit of 0.5 pmol
and the limited extent of the indeterminacy introduced
(55.5 pg) per sample, two-fold less sensitive than our
by small variations in eye size, as given by facet number
method, and lacks complete separation between the
(Krafka, 1920). Compared with existing HPLC meth-
histamine peak and neighboring peaks.
ods for histamine determination, our method is rapid,
The lack of a detectable histamine peak in the null
easy, of sufficient sensitivity, and adapted specifically to
mutant hdcJK910 confirms two critical pieces of evi-
work with these flies. Given the current interest in the
dence. First, it provides good evidence that the detected
visual system of Drosophila (Heisenberg and Wolf,
peak in Drosophila tissue lacking the synthetic enzyme
1984; Zipursky and Rubin, 1994; Ranganathan et al.,
for histamine is, indeed, that of histamine itself. Sec-
1995), and in transmitter systems (Na¨ssel, 1991; Restifo
ond, the lack of a detectable histamine peak means that
and White, 1990) in this genetically manipulable organ-
such a peak in wild-type tissue does not obscure peaks
ism, we developed an HPLC method specifically for
with similar retention times corresponding to other
histamine, because this is the reported transmitter for
substances in the sample. Melzig et al. (1998) have
fly photoreceptors (Hardie, 1987; Sarthy, 1991).
shown by immunocytochemical means that hdcJK910 can
Most older methods for histamine assay require the
take up exogenous histamine into the photoreceptors.
prior purification of this amine from biological samples
The fact that we failed to find a measurable histamine
before injecting the sample into the HPLC system
peak in hdcJK910 therefore implies that our flies were
(Davis et al., 1979; Yamatodani et al., 1982; Harsing et
unable to concentrate exogenous histamine to a signifi-
al., 1986; Han and Vohra, 1991). Thus, although such
cant extent from their diet. This in turn suggests that
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
there is a low concentration of histamine in our
the fluorescent or electrochemical detection of such
medium, as indeed was shown empirically. On the other
compounds (Simons and Johnson, 1978; Allison et al.,
hand, the presence of a small residual histamine peak in
1984). On the other hand, L-histidine, which does react
some chromatograms, close to the detection limit, could
with OPA-ME, is also undetectable in the range of
indicate either the uptake of exogenous histamine at
1 – 80 ng per sample (data not shown). Most probably
low concentrations from the medium, or possible resid-
L-histidine has a short retention time and disappears
ual expression of the hdc gene.
into the initial noise of the chromatograph. The same
Previous determinations of total histamine, using bio-
may also be true for other histamine-related com-
chemical methods (Elias and Evans, 1983) or HPLC
pounds which are not detectable with our method.
(Pirvola et al., 1988), report values for larger species of
Another proposed metabolite, carcinine, which is de-
insect, the locust and sphinx moth (Elias and Evans,
tected clearly in standard solutions, fails to give a
1983) and the cockroach (Periplaneta: Elias and Evans,
consistent peak in samples. Even so, the determination
1983; Blaberus: Pirvola et al., 1988). Most histamine
of histamine using a simple HPLC system equipped
occurs in the compound eyes (Elias and Evans, 1983),
with electrochemical detection is both easy and rapid in
about 150 times more than the amount occurring in the
the method we report that is especially adapted to
central brain (Pirvola et al., 1988). The determinations
reported here now show for the first time the accuratecontent of histamine in the Drosophila head. Our resultssupport in quantitative terms previous immunocyto-
Acknowledgements
chemical studies (Pollack and Hofbauer, 1991; Sarthy,1991; Na¨ssel and Elekes, 1992) which show that most
This work was supported by grants from MRC
histamine-like immunoreactivity occurs in the fly’s vi-
(MOP-36453), NIH (EY-03592) and NSERC (A-
sual system, corresponding to the strong reduction in
0000065). I.A.M. is also supported by the Killam Trust
the content of this amine in eyeless sine oculis. Thus
most histamine is contained in the compound eye’sphotoreceptors and their synaptic terminals. There isalso an action of eyelessness on the interneurons of the
References
lamina and the second neuropile, the medulla (Mein-
Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, Ama-
ertzhagen and Hanson, 1993), however. These neurons
lack immunoreactivity to histamine (Pollack and Hof-
melanogaster. Science 2000;287:2185 – 95.
bauer, 1991), so that their loss in sine oculis ought not
Allison LA, Mayer GS, Shoup RE. o-Phtalaldehyde derivatives of
contribute to the loss of histamine in this mutant,
amines for high-speed liquid chromatography/electrochemistry.
which we interpret as resulting entirely from the loss of
Arnould JM. Biosynthesis and metabolism of histamine in the central
photoreceptors. The residual histamine content of the
nervous system of Carcinus maenas. Arch Int Physiol Biochim
head sine oculis flies is presumably attributable to that
located in the integumentary mechanoreceptors (Buch-
Arnould JM. Beta-alanylation a means for neutralization of his-
ner et al., 1993), the other photoreceptor systems of the
tamine in the central nervous system in Carcinus maenas. Can J
ocellus (Pollack and Hofbauer, 1991) and eyelet (Hof-
Physiol Pharmacol 1987;65:1898 – 902.
Battelle B-A, Calman BG, Grieco FD, Mleziva MB, Callaway JC,
bauer and Buchner, 1989; Yasuyama and Meinertzha-
Stuart AE. Histamine: a putative afferent neurotransmitter in
Limulus eyes. J Comp Neurol 1991;305:527 – 42.
Buchner E, Buchner S, Burg MG, Hofbauer A, Pak WL, Pollack I.
Metabolites of histamine in invertebrates are not yet
Histamine is a major mechanosensory neurotransmitter candidate
well identified. The substances already proposed as the
in Drosophila melanogaster. Cell Tissue Res 1993;273:119 – 25.
Burg MG, Sarthy PV, Koliantz G, Pak WL. Genetic and molecular
main products of histamine metabolism in arthropods
identification of a Drosophila histidine decarboxylase gene re-
are: N-acetyl-histamine and imidazole-4-acetic acid
(Elias and Evans, 1983; Sarthy, 1991) or b-alanyl his-
tamine (carcinine: Arnould, 1985, 1987). In our method
Callaway JC, Stuart AE. Biochemical and physiological evidence that
neither N-acetyl histamine nor 4-imidazoleacetic acid
histamine is the transmitter of barnacle photoreceptors. Vis Neu-rosci 1989;3:311 – 25.
are detectable. Other methods for histamine determina-
Davis TP, Gehrke CW Jr, Gehrke CW, Cunningham TD, Kuo KC,
tion (Scofitsch et al., 1981; Harsing et al., 1986) also fail
Gerhardt KO. High-performance liquid chromatographic analysis
to measure acidic metabolites of histamine (imidazole-
of biogenic amines in biological materials as o-phtalaldehyde
4-acetic acid and 1-methyl-4-imidazoleacetic acid) after
derivatives. J Chromatogr 1979;162:293 – 310.
OPA-ME derivatization, using either fluorescence or
Elias MS, Evans PD. Histamine in the insect nervous system: distri-
bution, synthesis and metabolism. J Neurochem 1983;41:562 – 8.
electrochemical detection. It is therefore possible that
Elias MS, Lummis SR, Evans PD. 3[H] Mepyramine binding sites in
these substances do not react with OPA-ME, a reaction
the optic lobe of the locust: autoradiographic and pharmacologi-
which is specific for primary amines to improve either
cal studies. Brain Res 1984;294:359 – 62. J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Han XQ, Vohra MM. A sensitive method for simultaneous determina-
Pollack I, Hofbauer A. Histamine-like immunoreactivity in the visual
tion of histamine and noradrenaline with high-performance liquid
system and brain of Drosophila melanogaster. Cell Tiss Res
Ranganathan R, Malicki DM, Zuker CS. Signal transduction in
Hardie RC. Is histamine a neurotransmitter in insect photoreceptors?
Drosophila photoreceptors. Ann Rev Neurosci 1995;18:283 – 317.
J Comp Physiol 1987;A161:201 – 13.
Restifo LL, White K. Molecular and genetic approaches to neuro-
Hardie RC. Effects of antagonists on putative histamine receptors in
transmitter and neuromodulator systems in Drosophila. Adv Insect
the first visual neuropile of the housefly (Musca domestica). J Exp
Roeder T. Histamine H -receptor-like binding sites in the locust
Hardie RC. A histamine-activated chloride channel involved in neuro-
nervous tissue. Neurosci Lett 1990;116:331 – 5.
transmission at a photoreceptor synapse. Nature 1989;339:704 – 6.
Saito K, Horie M, Nakazawa H. Determination of urinary excretion
Harsing LG, Nagashima H, Vizi ES, Duncalf D. Electrochemical
of histamine and 1-methylhistamine by liquid chromatography. J
determination of histamine derivatized with o-phtalaldehyde and
2-mercaptoethanol. J Chromatogr 1986;383:19 – 26.
Sarthy PV. Histamine: a neurotransmitter candidate for Drosophila
Heisenberg M, Wolf R. Vision in Drosophila. Berlin, Heidelberg, New
photoreceptors. J Neurochem 1991;57:1757 – 68.
Schlemermeyer E, Schu¨tte M, Ammermu¨ller J. Immunohistochemical
Hofbauer A, Buchner E. Does Drosophila have seven eyes? Naturwis-
and electrophysiological evidence that histamine is a photorecep-
tor transmitter in locust ocellar retina. Invest Ophthalm Vis Sci
Jensen TB, Marley PD. Development of an assay for histamine using
automated high-performance liquid chromatography with electro-
Scofitsch G, Saria A, Holzer P, Lembeck F. Histamine in tissue:
chemical detection. J Chromatogr B 1995;670:199 – 207.
determination by high-performance liquid chromatography after
Krafka J. The effect of temperature upon facet number in the bar-eyed
condensation with ø-phtaldialdehyde. J Chromatogr 1981;226:53 –
mutant of Drosophila. Part I. J Gen Physiol 1920;2:409 – 32.
Lewis EB. A new standard food medium. Drosophila Info Serv
Simmons PJ, Hardie RC. Evidence that histamine is a neurotransmit-
ter of photoreceptors in the locust ocellus. J Exp Biol
Lundquist CT, Baines RA, Bacon JP. Evidence that histamine is a
neurotransmitter in an insect extraocular pathway. J Exp Biol
Simons SS Jr, Johnson DF. Reaction of 0-phtalaldehyde and thiols
with primary amines: fluorescence properties of 1-alkyl (and aryl)
Meinertzhagen IA, Hanson TE. The development of the optic lobe.
In: Bate M, Martinez Arias A (eds.). The Development of
thio-2-alkylisoindoles. Anal Biochem 1978;90:705 – 25. Drosophila melanogaster. Cold Spring Harbor Laboratory Press,
Stein C, Weinreich D. Metabolism of histamine in the CNS of Aplysiacalifornica: cellular distribution of g-glutamylhistamine synthetase.
Melzig J, Burg M, Gruhn M, Pak WL, Buchner E. Selective histamine
Comp Biochem Physiol 1983;74C:79 – 83.
uptake rescues photo- and mechanoreceptor function of histidine
Stuart AE. From fruit flies to barnacles, histamine is the neurotrans-
mitter of arthropod photoreceptors. Neuron 1999;22:431 – 3.
Yamatodani A, Fukuda H, Wada H. High-performance liquid chro-
Na¨ssel DR. Neurotransmitters and neuromodulators in the insect
matographic determination of plasma and brain histamine without
visual system. Progr Neurobiol 1991;37:179 – 254.
previous purification of biological samples: cation-exchange chro-
Na¨ssel DR, Elekes K. Aminergic neurons in the brain of blowflies and
matography coupled with post-column derivatization fluorometry. Drosophila: dopamine- and tyrosine hydroxylase-immunoreactive
neurons and their relationship with putative histaminergic neu-
Yamatodani A, Maeyama K, Watanabe T, Wada H, Kitamura Y.
rons. Cell Tissue Res 1992;267:147 – 67.
Tissue distribution of histamine in a mutant mouse deficient in
Na¨ssel DR, Holmqvist MH, Hardie RC, Ha˚kanson R, Sundler F.
mast cells: clear evidence for the presence of non-mast-cell his-
Histamine-like immunoreactivity in photoreceptors of the com-
tamine. Biochem Pharmacol 1982;31:305 – 9.
pound eyes and ocelli of the flies Calliphora eythrocephala and
Yasuyama K, Meinertzhagen IA. Extraretinal photorecptors at the
Musca domestica. Cell Tiss Res 1988;253:639 – 46.
compounds eye’s posterior margin in Drosophial melanogaster. J
Pirvola U, Tuomisto L, Yamatodani A, Panula P. Distribution of
histamine in the cockroach brain and visual system: an immunocy-
Zipursky SL, Rubin GM. Determination of neuronal cell fate: Les-
tochemical and biochemical study. J Comp Neurol 1988;
sons from the R7 neuron of Drosophila. Ann Rev Neurosci
Recommended procedure The caloric test General forward This document presents a Recommended Procedure by the British Society of Audiology (BSA). A Recommended Procedure provides a reference standard for the conduct of an audiological intervention that represents, to the best knowledge of the BSA, the evidence-base and consensus on good practice given the stated methodology and scope
Mid Back Complaints Today’s Date: _____/_____/_____ Name:_________________________________________________ Circle the areas on your body where you feel the described sensations, and mark with the appropriate letter(s). For Office Use Only: __________________________________________________ __________________________________________________ __________________________________________

 Journal of Neuroscience Methods 101 (2000) 141 – 148
The determination of histamine in the Drosophila head
J. Borycz a,*, M. Vohra c, G. Tokarczyk a, I.A. Meinertzhagen a,b
a Life Sciences Centre, Dalhousie Uni6ersity, Halifax, NS, Canada B3H 4J1
b Neuroscience Institute, Dalhousie Uni6ersity, Halifax, NS, Canada
c Department of Pharmacology, Sir Charles Tupper Building, Dalhousie Uni6ersity Medical School, 5859 Uni6ersity A6enue, Halifax,
NS, Canada B3H 4H7
Received 28 March 2000; received in revised form 31 May 2000; accepted 2 June 2000
Abstract
Journal of Neuroscience Methods 101 (2000) 141 – 148
The determination of histamine in the Drosophila head
J. Borycz a,*, M. Vohra c, G. Tokarczyk a, I.A. Meinertzhagen a,b
a Life Sciences Centre, Dalhousie Uni6ersity, Halifax, NS, Canada B3H 4J1
b Neuroscience Institute, Dalhousie Uni6ersity, Halifax, NS, Canada
c Department of Pharmacology, Sir Charles Tupper Building, Dalhousie Uni6ersity Medical School, 5859 Uni6ersity A6enue, Halifax,
NS, Canada B3H 4H7
Received 28 March 2000; received in revised form 31 May 2000; accepted 2 June 2000
Abstract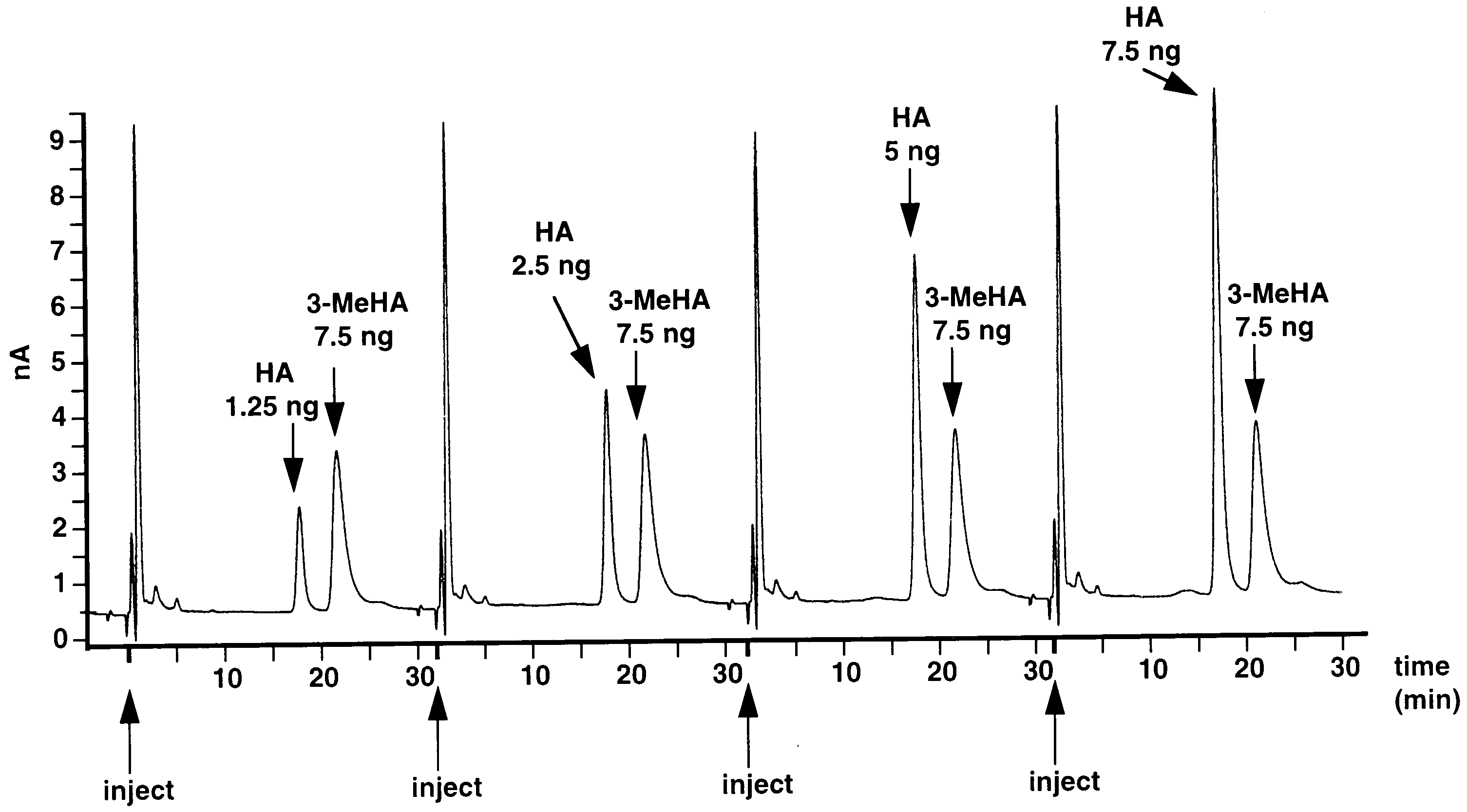
 J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Fig. 1. Detector response to increasing concentrations of injected standard histamine (HA), after condensation with OPA/ME. As an internalstandard 7.5 ng 3-methylhistamine (3-MeHA) was added to each sample. HPLC condition: 5 mm C18, 100 A, Nucleosil column (150×4.6 mm)coupled with a 5 mm C18, 100 A, Nucleosil guard column (10×4.6 mm); mobile phase: sodium acetate 100 mM; citric acid monohydrate 20 mM;1-octane-sulfonic acid, sodium salt 0.32 nM; Na EDTA 0.15 mM, acetonitrile 18% v/v and methanol 7% v/v; flow rate 1 ml/min.
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Fig. 1. Detector response to increasing concentrations of injected standard histamine (HA), after condensation with OPA/ME. As an internalstandard 7.5 ng 3-methylhistamine (3-MeHA) was added to each sample. HPLC condition: 5 mm C18, 100 A, Nucleosil column (150×4.6 mm)coupled with a 5 mm C18, 100 A, Nucleosil guard column (10×4.6 mm); mobile phase: sodium acetate 100 mM; citric acid monohydrate 20 mM;1-octane-sulfonic acid, sodium salt 0.32 nM; Na EDTA 0.15 mM, acetonitrile 18% v/v and methanol 7% v/v; flow rate 1 ml/min.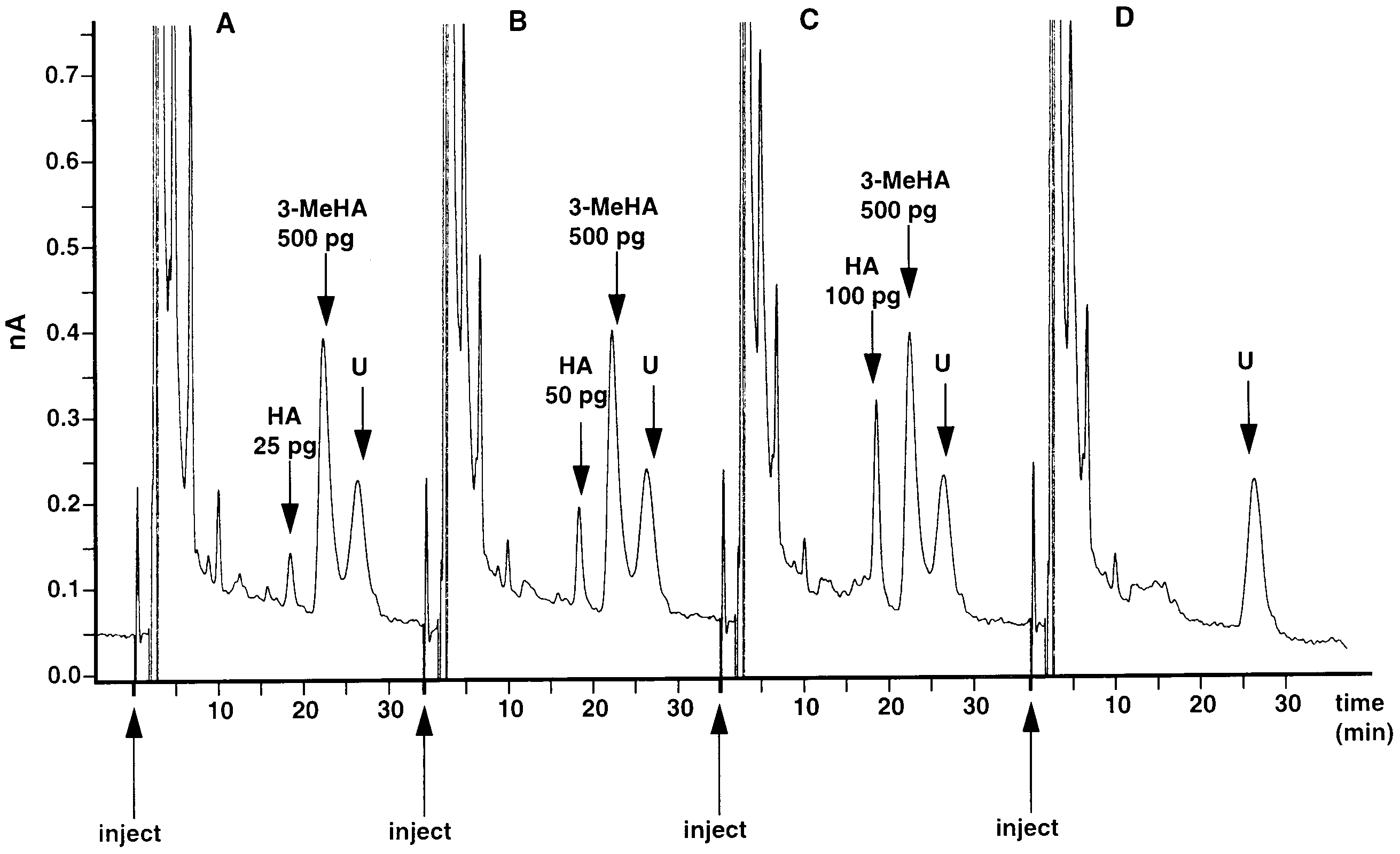
 J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Fig. 3. Lower limit of detection for histamine (HA). A,B,C: three chromatographs for increasing amounts of histamine (25, 50, 100 pg: arrows)which, as here, sometimes occur against a drifting baseline. As an internal standard, 3-methylhistamine (3-MeHA) 500 pg is added to each sample.
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
Fig. 3. Lower limit of detection for histamine (HA). A,B,C: three chromatographs for increasing amounts of histamine (25, 50, 100 pg: arrows)which, as here, sometimes occur against a drifting baseline. As an internal standard, 3-methylhistamine (3-MeHA) 500 pg is added to each sample.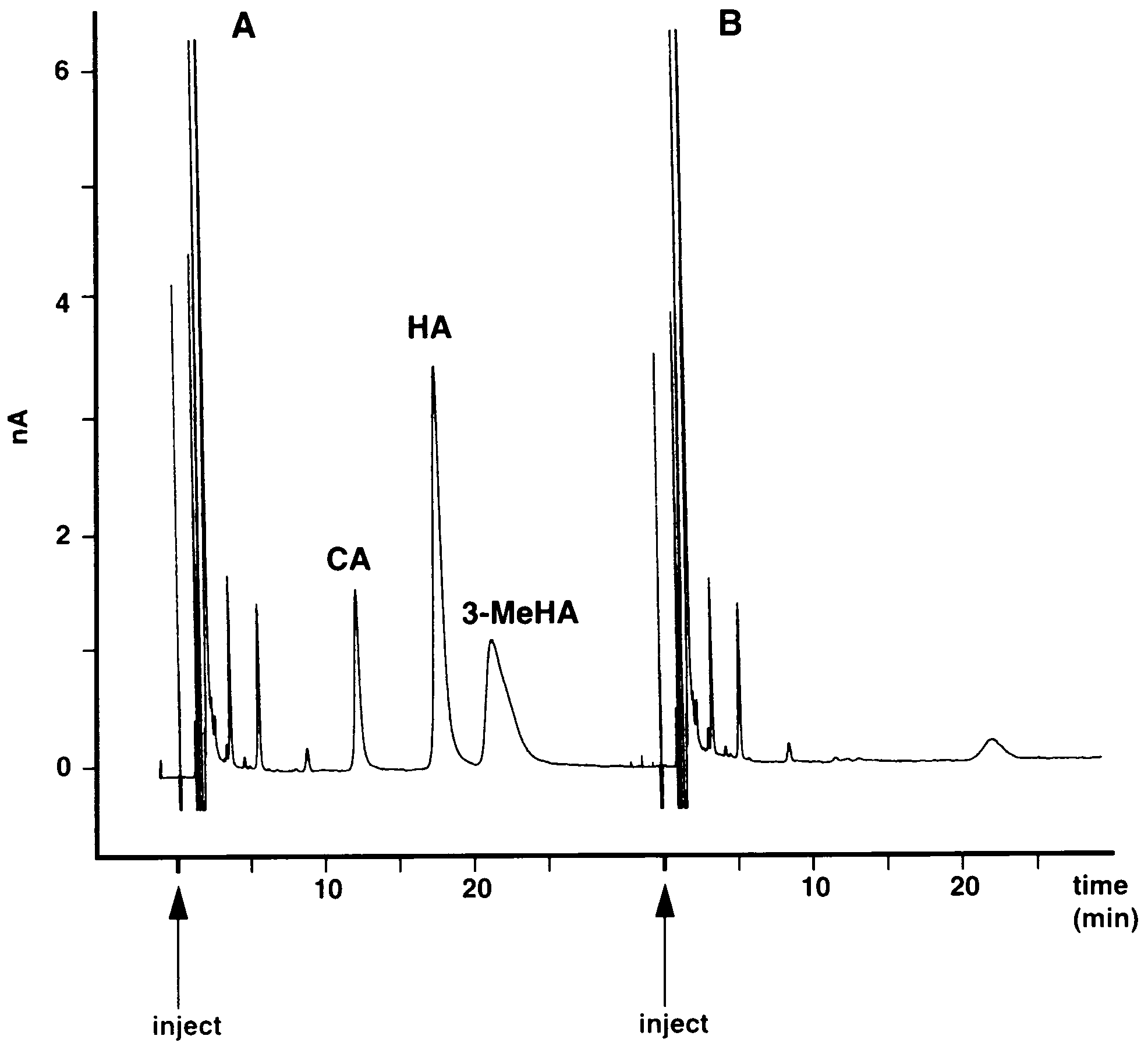
 J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
hdcJK910, head extracts of which lacked a clear peak atthe same retention time as the histamine peak in wild-type extracts (Fig. 6). Although some of the hdcJK910samples did in fact show a visible histamine peak, thiswas extremely reduced and never greater than the peakgiven by a 100 pg histamine standard.
J. Borycz et al. / Journal of Neuroscience Methods 101 (2000) 141 – 148
hdcJK910, head extracts of which lacked a clear peak atthe same retention time as the histamine peak in wild-type extracts (Fig. 6). Although some of the hdcJK910samples did in fact show a visible histamine peak, thiswas extremely reduced and never greater than the peakgiven by a 100 pg histamine standard.